



東工大ニュース
東工大ニュース
![]()
公開日:2014.09.24
東京工業大学大学院生命理工学研究科の木村宏教授と米国コロラド州立大学のスタセビッチ・ティモシー助教授(元・大阪大学大学院生命機能研究科)らの共同研究グループは、遺伝子の活性化の仕組みを生きた細胞の観察により明らかにした。ヒストンH3アセチル化[用語1] と転写を行う活性化型のRNAポリメラーゼII[用語2] を同時に生細胞で可視化し、数理モデルと合わせた解析により、ヒストンH3アセチル化が転写因子[用語3] のDNAへの結合と転写の伸長反応の両方に働くことを突き止めた。
細胞内でDNAと複合体を形成するヒストンたんぱく質の翻訳後修飾[用語4] は、遺伝子の抑制や活性化に働くと考えられている。中でもヒストンH3のアセチル化は、遺伝子活性化の目印として知られていたが、実際に細胞内でどのように働くのかは分かっていなかった。
共同研究には九州大学、かずさDNA研究所、モノクローナル抗体研究所などが参加した。成果は21日18時(英国時間)に英科学誌「ネイチャー(Nature)」オンライン速報に掲載される。
遺伝子は細胞の形や機能を担うたんぱく質の設計図であり、遺伝子が働くためには、その情報が必要に応じて正確にmRNAとして読み取られる必要がある。RNAポリメラーゼIIは、mRNAの転写を行う酵素であり、転写因子や「エピジェネティクス」制御[用語5] などにより、その遺伝子への結合や転写開始、転写伸長などのさまざまな過程が調節される(図1)。
ヒストンの翻訳後修飾の中でも、特にヒストンH3のアセチル化修飾は遺伝子活性化の目印として働くと考えられており(図1)、発生や分化、iPS細胞(人工多能性幹細胞)の形成過程で大きく変動することが知られている。
しかし、細胞内において、ヒストンH3アセチル化が、RNAポリメラーゼIIによる転写の調節に実際にどのように働くのかについての知見はほとんど得られていなかった。
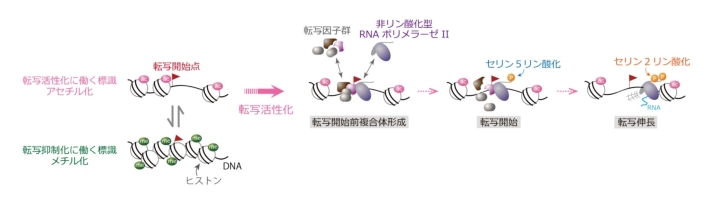
図1. ヒストン修飾と転写
DNAと複合体を形成するヒストンたんぱく質は様々な翻訳後修飾を受けるが、一般に、アセチル化は遺伝子の転写活性化に、メチル化は転写抑制に働く。転写には、RNAポリメラーゼと転写因子の遺伝子への結合が必要である。最初にDNAに結合し、転写開始前複合体を形成するのはリン酸化されていないRNAポリメラーゼII(非リン酸化型)であるが、転写開始時には繰り返しアミノ酸配列中の5番目のセリンが、転写伸長時には2番目のセリンがリン酸化される。従って、これらのリン酸化を特異的に検出することで、転写状態が分かる。
木村教授らは、これまで修飾特異的抗体を改変した蛍光プローブ[用語6] を用いて生細胞や生体内でヒストンの翻訳後修飾を計測する方法を開発してきた。今回、その方法を遺伝子の転写を行う蛋白質である活性化型RNAポリメラーゼIIの指標となるリン酸化修飾に適用し、転写の開始と伸長を生細胞で計測することに成功した。転写活性化のモデル系として、ステロイド系ホルモンである糖質コルチコイド[用語7] で誘導される遺伝子を用いた。
ホルモン誘導性遺伝子の繰り返し配列(遺伝子アレイ)を持つ細胞に、ヒストンH3修飾や活性化型RNAポリメラーゼIIに特異的に結合する蛍光プローブを導入し、その活性化に伴うヒストン修飾とRNAポリメラーゼIIの動態を生細胞で計測した。ホルモンの添加により、転写因子がすみやかに核に移行し、遺伝子アレイ上に集積する。今回の解析により、RNAポリメラーゼIIによる転写が、転写因子の結合後、数分以内に起こることが示された(図2)。
また、生細胞観察結果の定量解析と数理モデルの併用により、一旦、転写を開始した RNA ポリメラーゼII(セリン5リン酸化型)は効率よく伸長反応(セリン2リン酸化型)に至ることが明らかとなった。
さらに、ホルモン添加前に高レベルのアセチル化状態にある遺伝子アレイでは、転写開始の効率は低レベルのアセチル化状態にある遺伝子アレイと変わらないものの、転写因子の結合と転写の伸長が促進されることが明らかとなった。これらのことから、ヒストンH3のアセチル化は、転写因子の結合と転写開始から伸長への移行の両方を促進すると考えられる(図3)。この考えは、モデルとして用いた遺伝子アレイのみならず、他の遺伝子群についての解析結果とも一致した。従って、アセチル化による転写伸長反応の促進は、遺伝子の転写制御における普遍的な現象であると考えられ、今回の研究は、生命現象の理解に対して重要な意義を持つと考えられる。
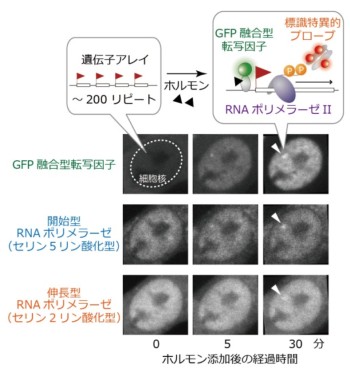
図2. ホルモン添加による遺伝子活性化
ホルモン誘導性遺伝子の繰り返し配列(遺伝子アレイ)を持つ細胞にホルモンを添加すると、遺伝子が活性化される。RNAポリメラーゼIIのリン酸化に特異的蛍光プローブを細胞に導入することで、この様子を生細胞で観察できる。ホルモンを添加すると、まず蛍光たんぱく質(GFP)融合型転写因子が細胞核内に移行し、遺伝子アレイに集積する。次いで、RNAポリメラーゼIIが遺伝子アレイに集積するが、転写開始型のセリン2リン酸化、転写伸長型のセリン5リン酸化が順次検出できる。
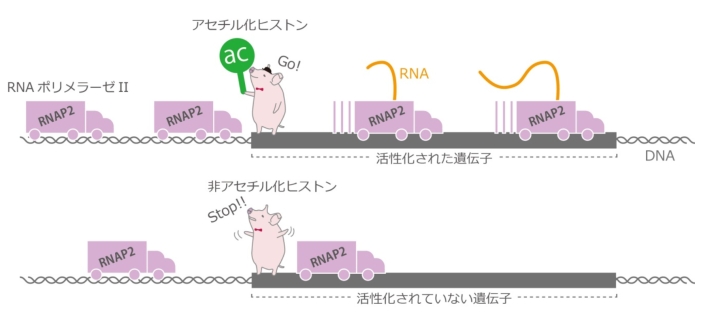
図3. ヒストンH3アセチル化の役割
ヒストンH3がアセチル化されていると、RNAポリメラーゼIIによる転写伸長反応が促進される。
今回着目したステロイド系ホルモン誘導性遺伝子は、あらかじめアセチル化されており、ホルモンに応答してすみやかに転写が起こるように準備された状態であると考えられる。今後、メチル化などの抑制的な修飾を持つ遺伝子が、発生や分化の過程でどのように活性化するのかを解明していく必要がある。
また最近、がんをはじめとした多くの疾患で、エピジェネティクス調節が異常になることが明らかにされはじめ、ヒストン翻訳後修飾を制御する因子を標的とする薬剤の開発が注目を集めている。本研究の知見や計測システムは、今後の薬剤開発にも大いに役立つと期待できる。
用語説明
[用語1] ヒストンH3アセチル化 : DNAは、ヒストンたんぱく質の周りを取り巻いて存在している。ヒストンには、H2A、H2B、H3、H4 の4種類存在するが、その中でもヒストンH3のアセチル化は転写の活性化に、メチル化は転写の抑制に働く。
[用語2] 活性化型のRNAポリメラーゼII : RNAポリメラーゼIIは、遺伝子の転写を行うたんぱく質であるが、その活性はリン酸化により制御される(図1)。DNAに結合するのは非リン酸化型であるが、転写開始時には繰り返しアミノ酸配列中の5番目のセリンがリン酸化され、転写伸長反応時には2番目のセリンがリン酸化される。
[用語3] 転写因子 : 遺伝子の転写開始や転写調節に働くたんぱく質で、DNAに直接結合して、転写を誘導するものが多い。
[用語4] 翻訳後修飾 : 蛋白質は細胞内で生合成された後、アセチル化、メチル化、リン酸化など様々な化学修飾を受ける。細胞内のほとんどの蛋白質は、これらの修飾により機能や活性が調節されている。
[用語5] エピジェネティクス制御 : 「エピジェネティクス」とは、DNA配列の変化を伴わずに起こる遺伝子発現の制御であり、DNAのメチル化やDNAと複合体を形成するたんぱく質であるヒストンの翻訳後修飾が重要な役割を果たしている。
[用語6] 蛍光プローブ : 生体内の現象を観察するための、蛍光を発する化合物や蛋白質などの小分子。細胞内で標的に特異的に結合するため、蛍光顕微鏡下で、標的分子の動態計測が可能になる。
[用語7] 糖質コルチコイド : 副腎皮質から分泌されるステロイド系のホルモン。糖代謝に関与し、生体内では血糖値の上昇作用や抗炎症作用などをもつ。
論文情報
掲載誌 : |
Nature |
論文タイトル : |
Regulation of RNA polymerase II activation by histone acetylation in single living cells |
著者 : |
Timothy J. Stasevich, Yoko Hayashi-Takanaka, Yuko Sato, Kazumitsu Maehara, Yasuyuki Ohkawa, Kumiko Sakata-Sogawa, Makio Tokunaga, Takahiro Nagase, Naohito Nozaki, James G. McNally, and Hiroshi Kimura
|
DOI : |
問い合わせ先
大学院生命理工学研究科 生体システム専攻
教授 木村 宏
Email: hkimura@bio.titech.ac.jp
TEL: 045-924-5742
