



東工大ニュース
東工大ニュース
![]()
公開日:2015.08.20
東京工業大学大学院生命理工学研究科の田中幹子准教授と鬼丸洸元大学院生(現CRG博士研究員)らの研究グループは、スペインCRG[用語1]のジェームズ・シャープ(James Sharpe)教授らと共同で、鰭(ひれ)から四肢への進化をもたらした要因の解明に成功した。軟骨魚類ハナカケトラザメ(Scyliorhinus canicula:以下、サメ)の鰭を解析し、サメ胚(個体発生の初期段階)の胸鰭は、付け根の部分が3つの骨からなる原始的な鰭の特徴を持つが、鰭から四肢への進化の過程で、前側と後側の位置価[用語2]をもつ領域のバランスが、後側寄りにシフトすることがわかった。
位置価のシフトの原因は、四肢の原基[用語3]の前側で発現するGli3 遺伝子[用語4]が、サメの胸鰭原基では後側で強く発現しているためであることが考えられた。さらに、Gli3 遺伝子の発現を変化させた原因となったゲノム配列を同定した。
そこで、サメ胚の鰭の位置価を人為的に後側寄りにシフトさせると、陸にあがる直前のティクターリク(Tiktaalik[用語5])の胸鰭のように、付け根の部分の骨が1つに融合し、先端の骨の数が減少していることがわかった。このことから、鰭から四肢への進化の過程では、Gli3 遺伝子の発現が変化したことで、前側と後側の位置価をもつ領域のバランスが、後側寄りにシフトすることが重要な原因のひとつであることが示された。
研究成果は8月18日に国際科学誌「eLife」[用語6]で公開された。
私たちの四肢は、原始的な魚類の鰭から進化したものである。原始的な魚類の鰭は、鰭の付け根の部分が3つまたはそれ以上の骨からなっており、この特徴を備えている鰭をもつ現存の生物としては、軟骨魚類があげられる(図1a)。一方、四肢の場合は、付け根の部分は1つの骨からなっている(図1a)。そこで、東京工業大学の田中准教授と鬼丸洸元大学院生らは軟骨魚類サメ胚を題材に、鰭が四肢へと進化したメカニズムを調べることにした。
四肢の原基では、前側(親指側)と後側の領域で様々な形態形成に関わる遺伝子が発現している(図1b)。軟骨魚類サメ胚の胸鰭の原基において、これらの遺伝子の発現パターンを調べてみると、前側と後側の領域のバランスが、四肢の原基よりも後側寄りになっていることがわかった(図1b)。これは、鰭から四肢への進化の過程で、前側と後側の位置価をもつ領域のバランスがシフトしたことを意味している。
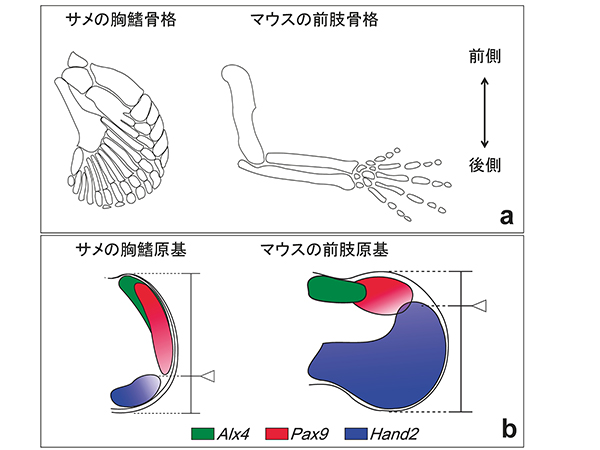
四肢の形成過程では、前側と後側の位置価をもつ領域のバランスは、四肢の原基の前側の広い領域で発現しているGli3 という遺伝子によって制御されている。そこで、サメ胚の胸鰭の原基において、Gli3 遺伝子の発現を調べたところ、サメの鰭では、Gli3 遺伝子が、四肢の原基とは異なり、後側で強く発現していることがわかった。
この特徴は、軟骨魚類全頭類[用語7]のゾウギンザメ胚の鰭でも確認されたことから、軟骨魚類の鰭で保存されている特徴と考えられた。さらに、Gli3 遺伝子の発現パターンの変化の原因について解析したところ、軟骨魚類から四肢動物への進化の過程においてGli3 遺伝子の発現を制御するゲノム配列が変化していることに起因していることを明らかにした。
これらの結果は、Gli3 遺伝子の発現パターンが変化したことにより、前側と後側の位置価をもつ領域のバランスがシフトして、鰭が四肢へと進化した可能性を示唆していた。そこで、サメの胸鰭の前側と後側の領域のバランスを人為的な方法(レチノイン酸という物質で処理する手法)でシフトさせ、後側化させることで、鰭を四肢へと変化させることができるか検証した。その結果、後側化させたサメの胸鰭は、絶滅した肉鰭類Tiktaalikでみられるような付け根に1つの骨をもつ四肢様の鰭に形を変化させることがわかった(図2)。
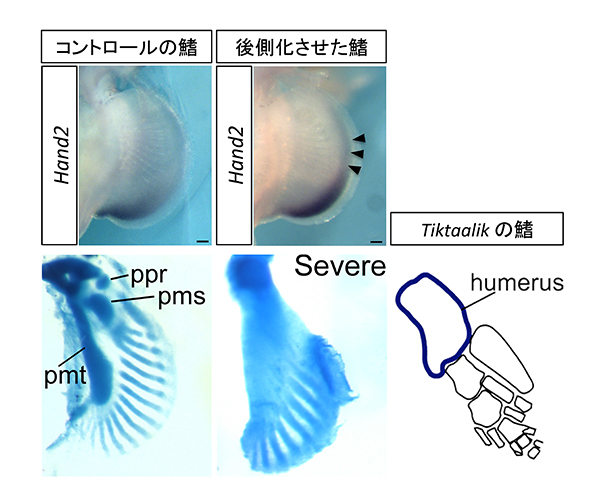
これらの結果から、鰭から四肢への進化の過程では、Gli3 遺伝子の発現パターンの変化により、前側と後側の位置価のバランスが少しずつシフトしていくことが、付け根の部分に1つの骨をもち、先端には5本の指をもつ四肢へと進化していく上で重要な要因の一つであったと考えられた(図3)。
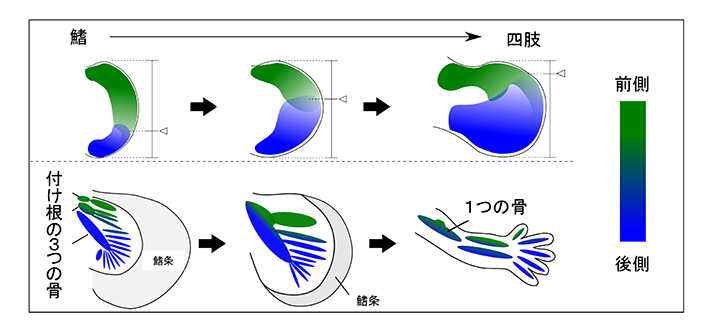
サメを題材にした今回の研究によって、鰭から四肢への進化が、前側と後側のバランスのシフトが一因となっていることを初めて示すことに成功した。Gli3 遺伝子の発現を制御するゲノム配列の変化が鍵であることを示したが、実際には、Gli3 とあわせて複数の因子が前側と後側のバランスのシフトに関わっていたと思われる。
また、鰭から四肢への進化の過程では、なぜ5本指になったのか(原始的な両生類は7-8本指あったと考えられている)などの問題は解明されていない。今後もサメを題材に、鰭から四肢への進化の過程で働くGli3 以外の因子や、その作用機序を明らかにしていくことで、鰭から四肢への進化の謎に迫りたい。
用語説明
[用語1] CRG : Center for Genomic Regulation スペイン・バルセロナにある生命科学の研究所
[用語2] 位置価 : 個々の細胞に与えられる分子的な番地表示のことで、体の中での相対的位置を示す。遺伝子発現レベルなどによって与えられる。
[用語3] 原基 : 将来ある器官になることに予定されてはいるが,まだ形態的・機能的には未分化の状態にある部分。
[用語4] Gli3 遺伝子 : 四肢では、前側と後側領域のバランスを決める鍵となる遺伝子。
[用語5] ティクターリク(Tiktaalik) : 3億年以上前に生息した絶滅肉鰭類で、四肢動物と多くの共通点を持つ。
[用語6] 「eLife」 : 生命科学・生命医学分野の一流の成果が発表されるオープンアクセス誌である(インパクトファクター 9.322)。
[用語7] 全頭類 : 軟骨魚類の現生種は板鰓類と全頭類に大きくわけられる。ハナカケトラザメ(文中の「サメ」)は板鰓類で、ゾウギンザメは全頭類。
論文情報
掲載誌 : |
eLife 2015;4:e07048. |
論文タイトル : |
A shift in anterior-posterior positional information underlies the fin-to-limb evolution. |
著者 : |
Koh Onimaru, Shigehiro Kuraku, Wataru Takagi, Susumu Hyodo, James Sharpe and Mikiko Tanaka
|
DOI : |
研究グループ
東京工業大学、Center for Genomic Regulation (CRG)、理化学研究所、東京大学
研究サポート
本成果は、文部科学省科学研究費補助金 基盤研究(B)、東京工業大学グローバルCOEプログラム「生命時空間ネットワーク」、日本学術振興会 日本-オーストラリア二国間交流事業、稲盛財団研究助成金、CREA、及び CRG のサポートを受けて得られた。
問い合わせ先
大学院生命理工学研究科生体システム専攻
准教授 田中幹子
Email : mitanaka@bio.titech.ac.jp
Tel : 03-5841-7251
東京工業大学 広報センター
Email : media@jim.titech.ac.jp
Tel : 03-5734-2975 / Fax : 03-5734-3661
