



東工大ニュース
東工大ニュース
![]()
公開日:2016.05.10
藻類の「眼」が正しく光を察知する機能を解明
―「眼」の色は細胞のレンズ効果を防ぐために必要だった―
東京工業大学 科学技術創成研究院 化学生命科学研究所の植木紀子研究員、井手隆広研究員(現・理研CDB研究員)、若林憲一准教授らの研究グループは、単細胞緑藻クラミドモナスが示す走光性(照射される光に反応して生物が移動する性質[用語1])の正と負が、眼点への色素集積を失った突然変異株では入れ替わることを発見した。
クラミドモナスは鞭毛[用語2]を使って水中を泳ぐ生物で、細胞の光反応行動の実験材料としてよく用いられる。クラミドモナス野生株のゲノムに対しランダム変異導入[用語3]を行って、「野生株と逆の走光性を示す突然変異株」を単離した。次世代シーケンサー[用語4]などによって、逆の走光性を示す原因となる遺伝子を同定したところ、カロテノイド色素[用語5]の生合成に関わる酵素に変異が入っていたことを突き止めた。
この色素は光受容体付近に存在し、これまでは細胞の光受容の指向性を高めるために存在すると考えられてきた。しかし色素を失った細胞がなぜ逆方向に泳ぐのか検証したところ、細胞が凸レンズの役割を果たして集光し、光源が光受容体の反対側にあるときのほうが光を強く感じていることを示す結果が得られた。
細胞レンズ効果は、透明な細胞ではその存在が知られていたが、緑色のクラミドモナスにおいてもはっきりとしたレンズ効果を持つことがわかった。
これらの結果から藻類は、自らの細胞が持つレンズ効果に打ち勝って正しい光源方向を察知するために、光受容体周辺にカロテノイド色素を濃縮・配列させたと考えられる。
この成果は、東京工業大学 科学技術創成研究院 化学生命科学研究所の久堀徹教授、田中寛教授、法政大学の廣野雅文教授、基礎生物学研究所の皆川純教授、重信秀治特任准教授らのグループとの共同研究によるもので、米国科学アカデミー紀要(PNAS)オンライン版に4月27日(米国東部時間)に掲載された。
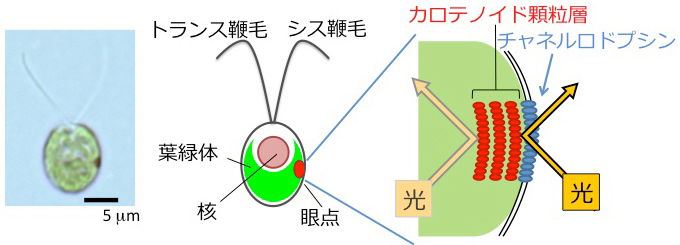
クラミドモナス(学名Chlamydomonas reinhardtii, 和名コナミドリムシ)は、淡水に棲む単細胞緑藻の一種である(図1)。2本の鞭毛を平泳ぎのように動かして水中を泳ぎまわり、かつ葉緑体で光合成するという、動物と植物の両方の特徴を合わせ持った生物である。
細胞に1つ「眼点」と呼ばれる光受容器官を持ち、光の指す方向を正確に認識する。光源方向に泳ぐ正の走光性と、逆に光源から逃げて泳ぐ負の走光性を適宜切り替えることで、生存に最適な光環境へ向かう。このような特徴をもつクラミドモナスは、鞭毛の運動調節、光合成、藻類の光環境適応行動など、広い研究分野で実験材料として用いられている。
光受容器官である眼点は、カロテノイド色素を豊富に含んだ顆粒が積層しているため、顕微鏡で観察すると赤い点のように見える(図1)。そのすぐ近傍の細胞膜に光受容タンパク質「チャネルロドプシン[用語6]」が存在する。このタンパク質は、光を受容すると開く陽イオンチャネルであり、流入した陽イオンがもとになる反応経路によって、細胞は光受容したあとの運動調節を開始する。
この「色素顆粒層」と「光受容タンパク質」がペアになっていることが、クラミドモナスの高指向性光受容を可能にしている。色素顆粒層は光をよく反射する性質をもつため、細胞の外側から来た光は増幅され、細胞の内側を通ってきた光は遮蔽されて受容体に届かない。クラミドモナスは、この「細胞の外から来た光しか感じない」というしくみよって、光の指す方向を正確に知るのである。
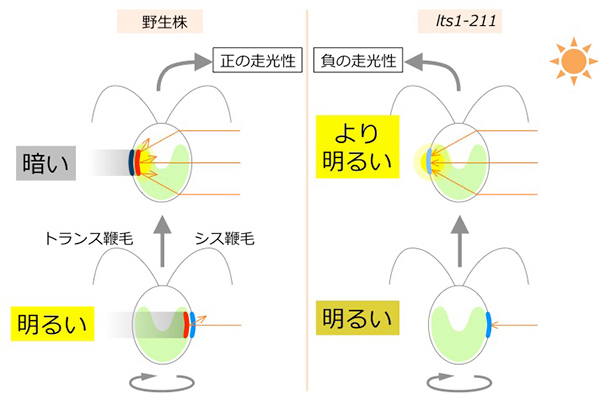
光受容後、2本の鞭毛(眼点に近いシス鞭毛と遠いトランス鞭毛に区別される)は打つバランスを変えることによって方向転換をする。光受容時にトランス鞭毛を強く打てば正の、シス鞭毛を強く打てば負の走光性を示す。若林准教授らは、数年前に活性酸素薬剤と活性酸素除去剤によって野生株の走光性を正または負にそれぞれ固定できることを見出したが、これらの薬剤の作動メカニズムはまだ不明である。
では、この眼点色素が失われたらどうなるのか。これまでは、「光が細胞の外からきたのか内から来たのか区別できなくなり、走光性を示さなくなる」と考えられてきた。
今回研究グループは走光性の正と負が決まる反応経路の同定を目指し、その経路に異常があると考えられる突然変異株の単離を試みた。野生株ゲノムにランダムに変異を導入し、得られた突然変異体群の中から、野生株と逆の走光性を示す新しい突然変異株を単離することに成功した。lts1-211と名付けた株は、野生株が正の走光性を示すときに負、負の走光性を示すときに正の走光性を示す(図2)。次世代シーケンサー[用語4]による全ゲノム解読などにより、lts1-211がカロテノイド生合成経路で機能する酵素に1アミノ酸置換変異をもつことがわかった。lts1-211細胞を詳細に顕微鏡観察すると、確かに眼点の色素が見られなかった(図2)。

眼点色素がないlts1-211はなぜ逆の走光性を示すのか。研究グループは「細胞レンズ効果」を仮定した。透明で球形の細胞が凸レンズのように振る舞って集光することは、ある種の菌類などで知られている。クラミドモナスにおいても、葉緑体色素を完全に失った透明な細胞では、レンズ効果を示す可能性は指摘されていた。しかし、それは直接には証明されておらず、また、葉緑体が正常で緑色の細胞がレンズ効果を示すか否かは議論が分かれていた。もしもレンズ効果を示すとすれば、光源が光受容体側でなく、その裏側にあるときのほうが、細胞は「より明るい」と感じるはずで、光源の方向を勘違いするだろう(図3)。
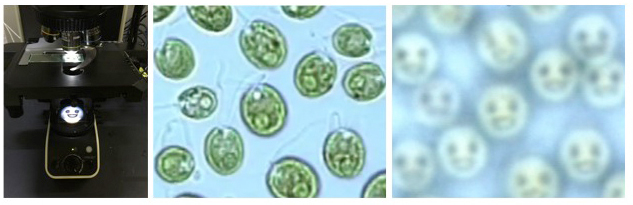
そこで、細胞がレンズ効果を持つか否かを確かめるために、顕微鏡光源付近に絵を置き、野生株細胞からフォーカスを少し上にずらして観察したところ、確かに絵が結像した(図4)。緑色の細胞がレンズ効果をもつことが初めてはっきりと示された。lts1-211が野生株と逆の走光性を示したのは、色素顆粒層の欠失により、細胞レンズ効果によって光源方向を勘違いしたせいだと考えられる。
つまり、眼点に存在している色素顆粒層は、「それがあることでより高い指向性が得られる」という補助的な役割ではなく、「それがなければ細胞のもつレンズ効果によって光源方向を逆側だと勘違いしてしまう」という、走光性の正と負を切り替える制御にとってなくてはならない構造であることが判明した。
今回得られた新しい突然変異株によって、走光性の正負の切り替えの反応経路の「根元」の部分が明らかになった。しかし、正常な色素顆粒層をもつ野生株が正負を切り替えるための、その後の反応経路の詳細はまだ明らかになっていない。今後、光受容以後の走光性の正負切り替え機構の解明を行っていきたい。そのような成果が得られれば、藻類の光受容システムの進化や、ヒトを含めた真核生物の鞭毛に共通した新たな鞭毛運動調節機構の理解につながると考えられる。
本研究は基礎生物学研究所共同利用研究「次世代DNAシーケンサー共同利用実験」のサポートを受けて実施された。
用語説明
[用語1] 走光性 : 生物が照射される光に反応して移動する性質。光源方向に近づく場合正の走光性、離れる場合負の走光性と呼ぶ。光走性(ひかりそうせい)とも。
[用語2] 鞭毛 : 真核生物細胞から生える毛状の運動する細胞小器官。細胞の推進力を生み出したり(精子など)、細胞の周囲に水流をつくったり(気管上皮など)、生体にとって重要な機能をもつ。ヒト体内には脳室、気管、輸卵管、精子などに運動性鞭毛・繊毛(鞭毛より短く本数が多いが、鞭毛と本質的に同じもの)が存在し、それらの運動異常によって生じる「原発性不動繊毛症候群」の研究にクラミドモナスは重要な役割を果たしている。
[用語3] ランダム変異導入 : 遺伝子組換え技術を使ってゲノムDNA上のある特定の塩基配列だけを狙って変異を導入する「部位特異的変異導入」に対して、紫外線照射などを使ってDNAに無作為的に変異を導入する手法。
[用語4] 次世代シーケンサー(第2世代シーケンサー) : 100塩基長程度に短く切断されたDNA断片の塩基配列を同時並行的に数千万以上決定することで、一度の大量の塩基配列データを取得するDNAシーケンサー。
[用語5] カロテノイド : カロチノイドとも。黄~赤色の天然色素。クラミドモナス眼点には主としてβカロテンが含まれる。
[用語6] チャネルロドプシン(ChR) : 光を感受すると構造変化してイオンチャネルとして機能する膜タンパク質。クラミドモナスにはChR1とChR2の2つがある。このうちChR2は、マウスなどの特定の神経細胞に発現させ、光照射によって興奮させることで神経活動と個体の行動の連関を研究する「光遺伝学」と呼ばれる技術に応用されている。
論文情報
掲載誌 : |
Proceedings of the National Academy of Sciences of the United States of America |
論文タイトル : |
Eyespot-dependent determination of the phototactic sign in Chlamydomonas reinhardtii |
著者 : |
*Noriko Ueki , *Takahiro Ide, Shota Mochiji, Yuki Kobayashi, Ryutaro Tokutsu, Norikazu Ohnishi, Katsushi Yamaguchi, Shuji Shigenobu, Kan Tanaka, Jun Minagawa, Toru Hisabori, Masafumi Hirono, and Ken-ichi Wakabayashi (*共同筆頭著者)
|
DOI : |
問い合わせ先
科学技術創成研究院 化学生命科学研究所
准教授 若林憲一
Email : wakaba@res.titech.ac.jp
Tel : 045-924-5235 / Fax : 045-924-5268
法政大学 生命科学部
教授 廣野雅文
Email : hirono@hosei.ac.jp
Tel : 042-387-6132
基礎生物学研究所 生物機能解析センター
特任准教授 重信秀治
Email : shige@nibb.ac.jp
Tel : 0564-55-7670 / Fax : 0564-55-7669
取材申し込み先
東京工業大学 広報センター
Email : media@jim.titech.ac.jp
Tel : 03-5734-2975 / Fax : 03-5734-3661
法政大学 広報課
Email : koho@hosei.ac.jp
Tel : 03-3264-9240 / Fax : 03-3264-9639
