



東工大ニュース
東工大ニュース
![]()
公開日:2018.01.18

東京工業大学 科学技術創成研究院 化学生命科学研究所の植木紀子研究員(現・ニューヨーク市立大学ブルックリン校上級研究員・ロックフェラー大学客員研究員)と若林憲一准教授は、多細胞緑藻であるボルボックスが、走光性[用語2]や光驚動反応[用語3]などの光に対する行動を示すために、球形の体の前端部から後端部にかけて鞭毛の性質を変えていることを発見した。
ボルボックスは鞭毛を使って水中を泳ぐ生物で、近縁の単細胞緑藻クラミドモナスに似た祖先生物の多細胞化によって進化したと考えられている。
約2億年前という比較的「最近」分岐したことや、祖先種に近いクラミドモナスが現存していることから、ボルボックスは多細胞化進化の研究の良い材料になっている。ボルボックスの約5,000個の細胞間には情報のやりとりが無いにも関わらず、個体として調和のとれた光行動を示す。
今回、この原理を明らかにするために鞭毛の性質を探ったところ、鞭毛は細胞の光受容に伴うCa2+(カルシウムイオン)の流入によって運動方向を変化させること、さらに、球のような体の前端部から後端部にかけて、その方向変化の角度が180度から0度まで変化することを突き止めた。ボルボックスは、個体前後で鞭毛の機能を変化させることで、前半球を舵取り、後半球を推進専門にと役割分担させ、巨大な体でも効率的な光行動を示すと考えられる。
この手法では、ボルボックスを界面活性剤処理で形態を保ったまま除膜する。この段階でボルボックスは死に、鞭毛が停止して動かなくなる。ここにATPを添加すると、個体は死んでいるのにも関わらず鞭毛が再び運動を開始して泳ぎだす。この手法は、単細胞生物や多細胞生物から取り出した器官でよく用いられる方法だが、多細胞生物個体まるごとを用いて成功したのは初めてであり、他の多細胞生物の運動メカニズムの検証にも適用できる可能性がある。さらに、鞭毛はヒトの体の様々な器官に生えており、各々は運動調節様式が異なる。この研究成果は、鞭毛運動の異常が原因であるヒトの疾患「原発性不動繊毛症候群」の研究に貢献すると期待される。
この成果は、米国科学アカデミー紀要(PNAS)オンライン版に1月8日に掲載された。
ボルボックス(和名:オオヒゲマワリ)は、淡水に棲む多細胞性の緑藻である(図1左上)。その最も大型のグループの1種ボルボックス・ルーセレティ(Volvox rousseletii)は、直径0.3~1 mm程度の球状の体の表面に約5,000個の細胞が一層の細胞層を形成している。1つ1つの細胞の構造は単細胞緑藻クラミドモナス(図1左下)によく似ており、各細胞は1つの光受容装置である眼点と、2本の鞭毛を持っている。ボルボックスには前後軸があり、約1万本の鞭毛は全て前から後ろに向かって運動する。これが原動力となってボルボックスは前進遊泳を行う。なお、「クラミドモナスが集まってボルボックスになる」、「ボルボックスがバラバラになってクラミドモナスになる」という話を聞くことがあるが、これは完全な誤りで、両者は近縁ではあるが別の生物だ。
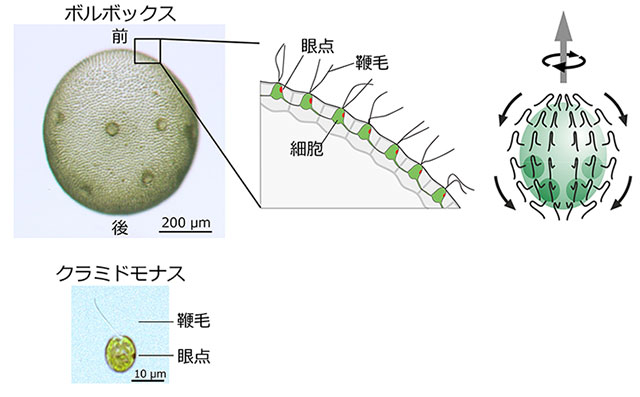
眼点は、赤い色素を豊富に含んだ顆粒が積層しているため赤い点のように見える。その直上の細胞膜に光受容タンパク質であるチャネルロドプシン[用語4]が存在する。これは光を受容すると開く陽イオンチャネルである。この光受容タンパク質と色素顆粒層がペアになっていることで、眼点は非常に指向性が高い光受容を行う(図2)。色素顆粒層は光をよく反射する性質を持つため、細胞の外側から来た光は増幅され、逆に細胞の内側を通ってきた光は遮蔽されて受容体に届かない。さらに、各鞭毛が打つ面は、個体の前後軸に対して少しだけ傾いているため、ボルボックスは遊泳する際にかならず進行方向後方からみて反時計周りに自転する(図1右)。各細胞は光源側を向いたときには光を感受し、個体が半回転して反対側を向いたときには光を感受しなくなる。このように、高指向性光受容と自転遊泳を組み合わせることによって、ボルボックスは光源方向を正確に認識する。
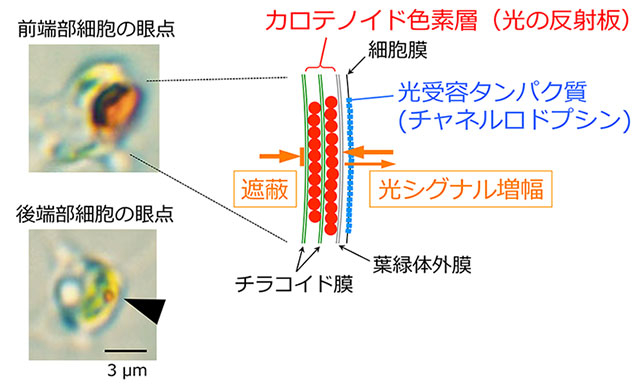
図2. 個体前端部付近の細胞(上)と後端部付近の細胞(下)の眼点と、眼点の模式図(右)
チャネルロドプシンとカロテノイド色素層の組み合わせにより、眼点は高い指向性をもった光受容を行う。眼点は前端部細胞では大きく、後端部細胞では小さい。
ボルボックスは各細胞の眼点で光を感受したのち、流入した陽イオンがもとになる反応経路によって鞭毛運動調節を行い、2つの光行動を見せる。1つは光驚動反応で、これは急に強い光を浴びたときに遊泳を停止する反応である。もう1つは走光性で、これは光源の方向に向かって、あるいは光源から逃げる方向に向かって遊泳する反応である。ボルボックスは、通常の条件下では主として光源に向かう正の走光性を見せる。これらは最適な光合成環境に移動するための生存戦略であると考えられている。
このとき、鞭毛はどのように動いているのか。以前、植木らは、ボルボックスが光を浴びたとき、前半球の細胞の鞭毛のみが運動方向を逆転させることを見出していた(Ueki et al., 2010 BMC Biol)。光驚動反応を示す際は前半球の鞭毛が前向きに打ち、後半球の鞭毛が後ろ向きに打つことで力が相殺されて個体の遊泳が止まる。走光性を示す際には、前半球かつ光源側の鞭毛だけが運動方向逆転を行うことで光源側と反対側で推進力に不均衡が生じ、光源方向に舵を切ることになる(図3)。
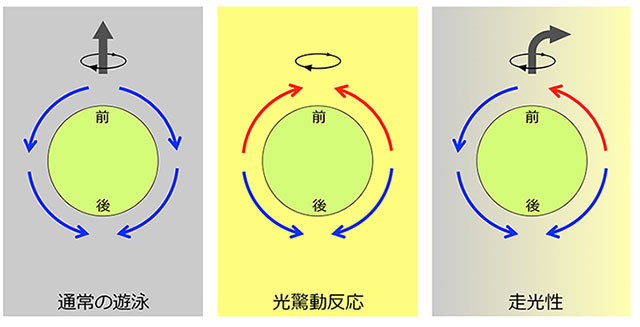
図3. ボルボックス鞭毛が起こす水流の方向
左:通常の遊泳時は全ての鞭毛が前から後ろへの水流を起こし、個体は自転しながら前進遊泳する。
中:急に強い光を浴びて光驚動反応を起こすとき、前半球の鞭毛は水流の方向を前向きに逆転させ、後半球の鞭毛は変わらず後ろ向きの水流を起こすため、個体は遊泳を停止し、その場で自転する。
右:右から光を浴びて正の走光性を示すとき、前半球の光源側の鞭毛だけが前向きの水流を起こし、他の部分は後ろ向きの水流を起こすため、個体の光源側とその反対側で推進力の不均衡が生じ、個体は右側に舵を切る。
この前後半球の鞭毛の光に対する応答性の違いは、これまで眼点の大きさの違いで説明されてきた(図2)。細胞がもつ眼点は、前端部に近いほど大きく、後端部に近いほど小さい。眼点が大きいほど光感受性が高いと考えられるため、後半球に比べて高い光感受性を持つ前半球の細胞の鞭毛だけが光に応答して運動方向を逆転する、という考え方だ。一方で、眼点の光感受の後に生じる鞭毛運動の方向逆転を起こす調節因子は不明だった。
今回研究グループは、鞭毛運動方向逆転の分子メカニズムを探るため、ボルボックスを用いた除膜モデルの試験管内での運動再活性化実験、通称“ゾンビ・ボルボックス法”の確立を試みた。まず界面活性剤処理によって細胞膜を溶解する。当然細胞は死ぬが、鞭毛の内部構造や運動するためのモータータンパク質はその形を留める。そして、ゾンビ・ボルボックスに生体エネルギー源であるATPを加えると、細胞は死んでいるが、鞭毛が再び運動を開始し、ボルボックスが泳ぎだす。これは1930年代にハンガリーのノーベル賞学者アルベルト・セントジェルジらによって行われたグリセリン筋の収縮実験に端を発する生物学の伝統的な手法だ。
この手法の最大のポイントはゾンビ・ボルボックスを入れる溶液の条件を自由に変えられることにある。この手法を用いた一連の実験では、筋肉の収縮や鞭毛運動のエネルギー源がATPであることが直接証明されてきた。ボルボックス鞭毛運動調節因子の第一候補はCa2+であり、この手法がボルボックスに適用できればCa2+の効果を直接確かめることができる。しかし、この手法は単細胞生物や多細胞生物から取り出した器官に対して適用されてきたもので、ボルボックスのような多細胞生物個体の場合、均一に界面活性剤処理をすることができなかったり、界面活性剤処理で形が崩れたりなど、技術的な高い障壁があった。
研究グループは、金魚すくいの要領でボルボックスを界面活性剤処理する手法を開発した(図4)。細胞ストレイナー(ふるい)の上で泳がせたボルボックスを持ち上げて、界面活性剤の入った溶液に漬け込む。これで個体全体が温和な条件で除膜される。ボルボックスの運動に関与する体細胞は球形の体の表面にあるので、これですべての体細胞は死に、鞭毛は運動を停止する。さらにふるいを持ち上げて界面活性剤のない溶液に漬け込み、ここにATPを加えると、ボルボックスは死んでいるものの、再び泳ぎだすことになる。研究グループはこのゾンビ・ボルボックス実験をさまざまなCa2+濃度条件下で行い、鞭毛運動を観察した。
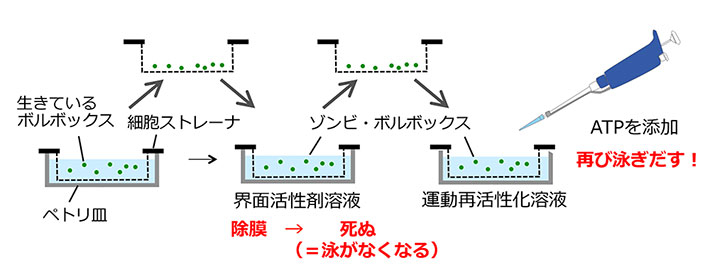
図4. ゾンビ・ボルボックス法の概要
細胞ストレーナ(目の細かいふるい)の上でボルボックスを泳がせる。ふるいを持ち上げて界面活性剤入りの溶液に漬け込むと、ボルボックスは除膜されて死に、泳がなくなる(ゾンビ・ボルボックスになる)。再びふるいを持ち上げて、界面活性剤のない溶液に漬け込み、ATPを添加すると、鞭毛が運動を再開し、ゾンビ・ボルボックスは死んでいるにも関わらず泳ぎだす。
Ca2+濃度が高い状態でゾンビ・ボルボックスにATPを加えると、前端部に近い鞭毛は運動方向をほぼ逆転させた(図5上)。これにより、これまで観察されていた鞭毛運動の方向逆転がCa2+によることが初めてはっきり示された。さらに興味深いことに、この運動方向変化の大きさには体の前後で違いがあった。前端部付近の細胞はほぼ180度の逆転、赤道面付近の細胞は90度程度、後端部付近の細胞は0度で、ほとんど逆転しなかった(図5下)。
ゾンビ・ボルボックスは細胞膜とともに光受容体を失っているので、光受容を行わない。そのため、この個体における細胞の位置に応じた鞭毛の運動性の違いは、純粋に鞭毛の性質の違いを反映している。つまり、これまで前半球と後半球で異なるのは眼点の大きさだけだと考えられてきたが、鞭毛の性質も異なることが今回初めて分かった。鞭毛打方向の「逆転」と考えられてきたことは、鞭毛打方向の個体の位置に応じた「回転」であった。これらの発見のためには、球状の個体の形を壊さずに、全体を温和に除膜するゾンビ・ボルボックス法の開発が不可欠だった。
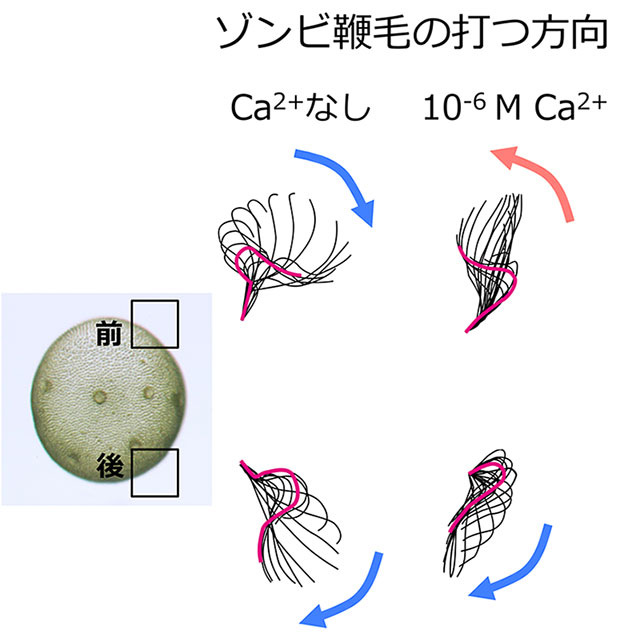
つまり、ボルボックスの細胞は、第一に眼点の大きさ、第二に鞭毛のCa2+応答性という2段構えの前後分化をしている。このことにより、前端部に近い細胞ほど個体に対してブレーキや舵取りを行う機能が高くなり、後端部に近い細胞ほど「何があっても前に進む」という推進力に特化していると言える。もしこの前後分化がなく、全ての細胞が同じ程度の光感受性と鞭毛打方向逆転能しかもっていなければ、横から光を受けた場合、個体はその場で回るだけで走光性は示せない。ボルボックス細胞の前後分化は、多細胞化によって巨大化した体で高い推進力を得ると同時に、単細胞緑藻のように機敏な光行動を行うために獲得した重要な機能であると考えられる。
ボルボックス目の緑藻は、単細胞性のクラミドモナスや約5,000細胞のボルボックス・ルーセレティだけでなく、中間の細胞数の多細胞種が多数現存している興味深い生物群だ。今回の手法を中間細胞数の藻類にも適用することで、多細胞化による光行動システムの変遷を探ることができる。
今回開発した“ゾンビ・ボルボックス法”は、さまざまな多細胞生物を温和に除膜モデル化するのに役立つ。藻類に限らず、多様な生物の運動システムの研究に貢献すると期待される。
Ca2+による鞭毛の運動調節は、ヒトを含めた多様な生物で見られる。ヒトの体内には、脳室、気管上皮、輸卵管上皮、精子など、さまざまな器官に鞭毛(繊毛)が生えている。これらの運動調節が異常になると、慢性呼吸器疾患や不妊症などのさまざまな疾患を誘起する。今後、この手法を用いて鞭毛運動調節の分子機構をさらに詳しく研究することで、鞭毛運動不全によるヒトの疾患である「原発性不動繊毛症候群」などの理解に貢献すると期待される。
本研究は科学研究費補助金(15H01206, 15H01314,16K14752)の支援を受けて行われました。
用語説明
[用語1] 鞭毛 : 真核生物細胞から生える毛状の運動する細胞小器官。精子のように細胞の推進力を生み出したり、気管上皮のように細胞の周囲に水流をつくったり、生体にとって重要な機能をもつ。ヒト体内には脳室、気管、輸卵管、精子などに鞭毛や繊毛(鞭毛より短く本数が多いが、鞭毛と本質的に同じ)が存在する。それらの運動異常によって生じる疾患は原発性不動繊毛症候群と呼ばれる。
[用語2] 走光性 : 生物が照射される光に反応して移動する性質。光源方向に近づく場合は正の走光性、離れる場合は負の走光性と呼ぶ。光走性(ひかりそうせい)と呼ばれることもある。
[用語3] 光驚動反応 : 生物が強い光強度変化に応答して、運動を止めたり、運動方向を逆転させたりする反応。光忌避反応と呼ばれることもある。
[用語4] チャネルロドプシン : 光を感受するとイオンを透過する膜タンパク質。マウスなどの特定の神経細胞に発現させ、光照射によって興奮させることで神経活動と個体の行動の連関を研究する「光遺伝学」と呼ばれる技術に応用されている。
論文情報
|
掲載誌 :
|
Proceedings of the National Academy of Sciences of the United States of America, in press.
|
|
論文タイトル :
|
Detergent-extracted Volvox model exhibits an anterior–posterior gradient in flagellar Ca2+ sensitivity
|
|
著者 :
|
Noriko Ueki and Ken-ichi Wakabayashi
|
|
DOI :
|
お問い合わせ先
東京工業大学 科学技術創成研究院 化学生命科学研究所
若林憲一 准教授
E-mail : wakaba@res.titech.ac.jp
Tel : 045-924-5235 / Fax : 045-924-5268
取材申し込み先
東京工業大学 広報・社会連携本部 広報・地域連携部門
E-mail : media@jim.titech.ac.jp
Tel : 03-5734-2975 / Fax : 03-5734-3661
