



東工大ニュース
東工大ニュース
![]()
公開日:2018.08.15
東京工業大学 科学技術創成研究院 化学生命科学研究所の吉田啓亮助教と久堀徹教授らの研究チームは、植物が夜間に光合成に関わる酵素をオフにするしくみを解明した。朝になって植物が光合成を始める際には、タンパク質分子の酸化と還元の切り替え機構である“レドックス制御[用語1]”の働きで光合成の糖代謝を担う酵素群が還元され、光合成機能が活性化される。この“オン”側のスイッチのしくみは古くから知られていたが、夜になったらどのようにして“オフ”にするのかは明らかにされていなかった。吉田助教らは、光合成の酵素群を酸化する(スイッチオフにする)分子機構を明らかにした。この研究成果は、植物がどのようにして夜間に光合成の糖代謝を抑えてエネルギー浪費を防ぐのか、そのしくみの一端を解明したものであり、環境適応型作物のデザインなど今後の応用展開のための重要な指針となると期待される。
研究成果は、2018年8月13日発行の米国科学アカデミー紀要 (Proc. Natl. Acad. Sci. USA)」電子版に掲載された。
植物の光合成は、地球規模で行われる壮大なエネルギー変換反応である。光合成は、私たちが呼吸するために必要な酸素を供給し、食糧となる炭水化物を生産している。植物の緑葉の細胞には、葉緑体と呼ばれる長径3~10 μmほどの細胞小器官があり、一連の光合成反応はこの細胞小器官内で行われている(図1)。さらに葉緑体の内部には、チラコイド膜とよばれる袋状に閉じた生体膜が積層している。チラコイド膜には、電子伝達反応を行うためのタンパク質分子装置が配置されている。電子伝達反応では、クロロフィルが捕集した光エネルギーを用いて水から電子を引き抜き、還元物質であるNADPHとエネルギー物質であるATPを合成する。また、この水の分解に伴って酸素が発生する。
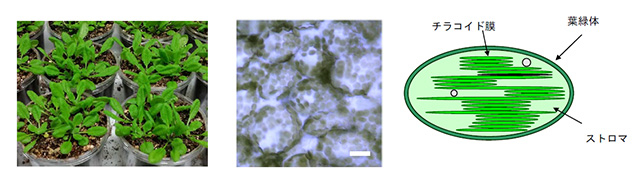
図1. シロイヌナズナの緑葉、表皮細胞(白バー:20 μm)と葉緑体模式図
葉緑体のストロマというゲル状の区画では、大気中から取り込まれた二酸化炭素を用いて有機物が合成される。この反応は13種類の糖代謝の酵素が連携して働く複雑な反応経路で行われており、発見者の名前を取ってカルビン・ベンソン回路と呼ばれている。カルビン・ベンソン回路が働くためには、チラコイド膜での電子伝達反応で作られたNADPHとATPが使われる。しかし、カルビン・ベンソン回路の酵素反応はいずれも光エネルギーを直接必要とはしない。そのため、光合成の研究初期はこの回路全体が“暗反応”と呼ばれ、明反応と呼ばれた電子伝達反応とは区別されていた。ところがその後、カルビン・ベンソン回路の4つの酵素は電子伝達反応が働くと酵素活性が高くなることがわかり、暗反応という定義が実態と合わなくなってしまった。植物に光が当たっている時に電子伝達系からこれらの酵素にシグナルを伝達し、酵素活性のスイッチをオンにするのが酸化還元(レドックス)制御システムである。
レドックス制御システムは、特定の酵素タンパク質が持っているジスルフィド結合の形成・開裂(酸化・還元)を生体内の酸化還元状態に応じて制御することで、その酵素の活性を調節する分子機構だ。この制御機構で中心的な役割を果たしているのが、チオレドキシン[用語2]という酸化還元タンパク質である。還元状態のチオレドキシンは、標的となる酵素が持っているジスルフィド結合を還元して開裂させることで構造変化を引き起こし、その酵素を通常は不活性型から活性型、つまりスイッチオンの状態にする。葉緑体では、電子伝達反応で得られた電子の一部をチオレドキシンが受け取ることでレドックス制御システムが働いている(図2)。
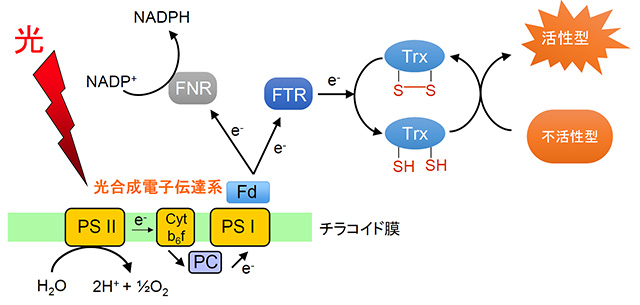
図2. 光合成電子伝達系から酵素への電子の受け渡し経路
PSII:光化学系II、Cyt b6f:シトクロムb6f複合体、PC:プラストシアニン、PSI:光化学系I、Fd :フェレドキシン、FNR:Fd-NADPレダクターゼ、FTR:Fd-チオレドキシンレダクターゼ、Trx :チオレドキシン
実際に植物体内で一日を通してどのようにレドックス制御システムが働いているのかを調べてみると、夜が明け光が強くなるのに応じて光合成に関わる複数の酵素が還元(スイッチオン)される(図3)。これが、上に述べた電子伝達反応から電子を受け取ったチオレドキシンの働きである。
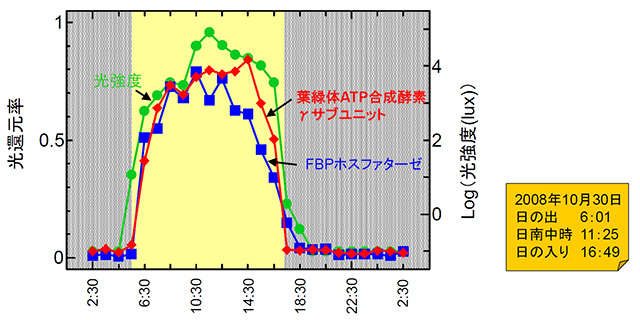
図3. 緑葉内の酵素の酸化還元状態の日周変化
ホウレンソウ緑葉の2つの酵素(ATP合成酵素とFBPホスファターゼ)の還元状態の変化をプロットしたもの。Konno, et al. (2013) Plant Cell Physiol. 53(4) : 626-634に掲載したデータより引用。
一方で、夕暮れになり光が弱くなると、今度はこれらの酵素が酸化(スイッチオフ)される。このとき、“何らかの酸化力”が働いていると思われるが、これに関わる分子の実態は長らく知られていなかった。2004年に国際光合成会議がモントリオールで開かれた際、レドックス討論会の座長を務めた久堀教授がこの問題を提起したときには、「チオレドキシンが酸化の過程も担っているのではないか」、「単純に酸素分子が直接酸化しているのだろう」などの議論があったが、誰も実証はできていなかった。
チオレドキシンは、活性部位に-WCGPC-というアミノ酸配列を持っている。2000年に緑色植物シロイヌナズナの全ゲノムのDNA塩基配列が解読され、植物は(オーソドックスな)チオレドキシン以外にも、アミノ酸配列がチオレドキシンに類似したタンパク質を複数持っていることがわかった。これらのタンパク質は、チオレドキシンと同様に酸化還元タンパク質として働くと予想された。吉田助教らは、その中で―WCRKC―というチオレドキシン活性部位に似たアミノ酸配列を持つ機能がわかっていないタンパク質に着目した。海外で行われた先行研究では、このタンパク質は “thioredoxin-like2” と名付けられていたので、それにならってTrxL2と呼ぶことにした。
TrxL2の細胞内局在や生化学的な性質を調べたところ、葉緑体のストロマに局在すること、還元力の伝達活性を持つこと、光合成の酵素群と相互作用することなどがわかった。これらは、既知の葉緑体のチオレドキシンの特徴と同じだ。ところがTrxL2は、葉緑体の酵素と物理的な相互作用はできるが、チオレドキシンのようにはそれらを還元することができなかった。
“酸化還元電位”は、還元力のやり取りのしやすさを定量的に評価するものさしである。そこで、吉田助教らはTrxL2の酸化還元電位を測定し、チオレドキシンと比較したところ、チオレドキシンよりも著しく高いことがわかった。この結果は、TrxL2は相手を還元するよりも、むしろ自身が還元されやすい(すなわち相手を酸化しやすい)ということを意味している。そこで、あらかじめ還元型にしておいた標的酵素と、酸化型のTrxL2を同じモル比で混合したところ、標的酵素は酸化され、それにつれてTrxL2は還元された。つまり、TrxL2は、チオレドキシンとは逆方向に還元力の受け渡しを行う酸化因子タンパク質なのだ。
しかし、TrxL2が葉緑体内に多く存在する光合成の酵素を酸化し続けるためには、一度受け取った還元力をさらに別の何かに渡す必要がある。吉田助教らは、その下流にある因子の同定にも成功した。TrxL2は、ほとんどのタンパク質に対して還元能を示さなかったが、例外的かつ極めて高い効率で2-システイン-ペルオキシレドキシン(2-Cys Prx)[用語3]を還元した。2-Cys Prxは、細胞にとって有害な活性酸素の消去に還元力を使う。すなわち、TrxL2は、最終的には活性酸素が持つ強い酸化力を利用して、持続的なタンパク質の酸化を行っているわけだ(図4)。
さらに、吉田助教らは、このようなタンパク質の酸化システムが実際に植物体内で働いていることを示すことにも成功した。2-Cys Prxを欠損したシロイヌナズナの変異株植物を用いて、明・暗に応答したタンパク質の還元・酸化の動態を野生株の植物と比較したところ、2-Cys Prxを欠いた植物では、明所から暗所に移されたときに光合成の酵素群の酸化が正常に進まず、同時にTrxL2に還元力が蓄積することがわかった。
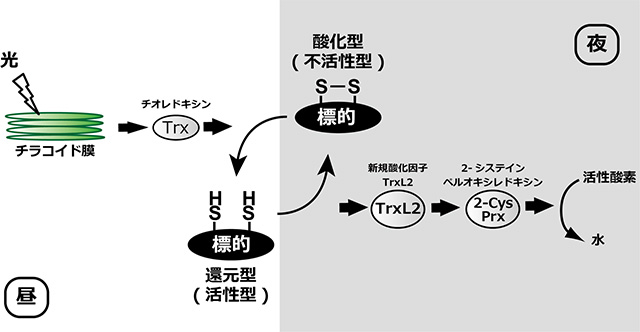
図4. 新たに解明したTrxL2による酸化経路
夜間、植物は光合成を行うことができない。本研究で明らかになったしくみは、光合成の糖代謝に関わる酵素群を夜に眠らせて無駄なエネルギー消費を抑えるための戦略といえる。TrxL2を介したタンパク質酸化システムは、植物葉緑体の祖先であるシアノバクテリアには見当たらない。すなわち植物は、昼夜サイクルを繰り返す陸上環境に適応するために、巧妙な酵素のオン・オフの切り替えスイッチを進化の過程で獲得したと考えられる。
本研究の成果は、現代の植物科学の基礎学術研究に新たなブレイクスルーをもたらすだけでなく、植物あるいは光合成微生物を利用した物質生産など、将来の応用展開にも重要な情報を提供するものである。
用語説明
[用語1] レドックス制御 : 酸化還元状態に応じて、タンパク質分子の持っているジスルフィド結合の形成・開裂などを制御することにより、そのタンパク質の酵素活性を調節する分子機構。タンパク質の翻訳後修飾のひとつ。
[用語2] チオレドキシン(Trx) : レドックス制御に中心的な役割を果たす酸化還元タンパク質。すべての生物が普遍的に持っている。-WCGPC-(-Trp-Cys-Gly-Pro-Cys-)というよく保存された活性部位モチーフを持ち、この2つのCysのチオール基の酸化還元によって還元力伝達を行う。植物葉緑体にはf, m, x, y, z型という5つの分子種が存在する。TrxL2の活性部位モチーフは、―WCRKC―(Trp-Cys-Arg-Lys-Cys)である。
[用語3] 2-システイン-ペルオキシレドキシン(2-Cys Prx) : チオレドキシンなどから還元力を受け取り過酸化水素を還元するタンパク質。生体を活性酸素種から守る抗酸化ストレスタンパク質として重要な役割を担っている。
論文情報
掲載誌 : |
Proc. Natl. Acad. Sci. USA 2018, in press |
論文タイトル : |
Thioredoxin-like2/2-Cys peroxiredoxin redox cascade supports oxidative thiol modulation in chloroplasts |
著者 : |
Keisuke Yoshida, Ayaka Hara, Kazunori Sugiura, Yuki Fukaya, and Toru Hisabori |
DOI : |
お問い合わせ先
東京工業大学 科学技術創成研究院 化学生命科学研究所 助教
吉田 啓亮
E-mail : yoshida.k.ao@m.titech.ac.jp
Tel : 045-924-5267 / Fax : 045-924-5268
東京工業大学 科学技術創成研究院 化学生命科学研究所 教授
久堀 徹
E-mail : thisabor@res.titech.ac.jp
Tel : 045-924-5234 / Fax : 045-924-5268
取材申し込み先
東京工業大学 広報・社会連携本部 広報・地域連携部門
Email : media@jim.titech.ac.jp
Tel : 03-5734-2975 / Fax : 03-5734-3661
