



東工大ニュース
東工大ニュース
![]()
公開日:2018.10.16
東京工業大学 科学技術創成研究院の岩﨑博史教授、伊藤健太郎研究員、黒川裕美子研究員、国立台湾大学の李弘文教授(Pro. Hung-Wen Li)、台湾国立中央研究院の冀宏源准教授(Assoc. Prof. Peter Chi)等からなる国際研究チームは、DNA相同組換えの中心的な反応である“DNA鎖交換反応”をつかさどるDNA―Rad51タンパク質[用語1]複合体形成制御のしくみを世界で初めて明らかにした。
相同組換えは、全ての生物で起きる生命現象で、傷ついたDNAの修復や遺伝的多様性を生み出すのに必須の働きをしている。相同組換えは、似た配列を持つ(このことを“相同”という)DNA鎖の交換反応が中心的な反応で、Rad51リコンビナーゼ[用語2]によって触媒される。
Rad51は、1本鎖DNAにらせん状に結合したフィラメント構造を作る。この構造体は、相同二重鎖DNAを検索してDNA鎖の交換を触媒する。しかしこの構造体は、かなり不安定であり、補助因子Swi5-Sfr1タンパク質複合体によって安定化される必要があるが、どのような分子機構でフィラメント構造が安定化するのか不明だった。
本研究では、フィラメント一分子をリアルタイムで観察することで、Rad51が1本鎖DNAへ結合・解離する過程を解析してSwi5-Sfr1複合体がRad51の解離を抑制しフィラメントの安定化を促進することを世界で初めて示した。
この成果は、10月8日(米国東部時間)付けの『Proceedings of the National Academy of Sciences of the USA(米国科学アカデミー紀要)電子版』に掲載された。
本研究では、Rad51が1本鎖のDNAと結合するとDNAを伸長させるという性質を利用して、(1) DNAの末端に微小なスチレンビーズを付加してビーズのブラウン運動を観察してDNAの伸長を検出する実験系、(2) DNA鎖を蛍光でラベルし、蛍光共鳴エネルギー移動 (Fluorescence resonance energy transfer: FRET)の原理を利用してDNAの伸長を検出する実験系を構築。1本鎖DNA上にRad51フィラメントが結合・解離する様子を一分子単位でリアルタイムに観察することに成功した。
これら2つの実験系を駆使し、様々な条件で解析した結果、まず、2~3分子のRad51が1本鎖DNA上に小さなフィラメント核を形成後、その核の末端にさらにRad51が結合してフィラメントが伸長していくことが分かった。さらに真核生物が共通して持つ複合体形成の促進因子であるSwi5-Sfr1タンパク質は、Rad51の1本鎖DNAからの解離を抑制することで、フィラメントを安定化することを明らかにした(図1)。
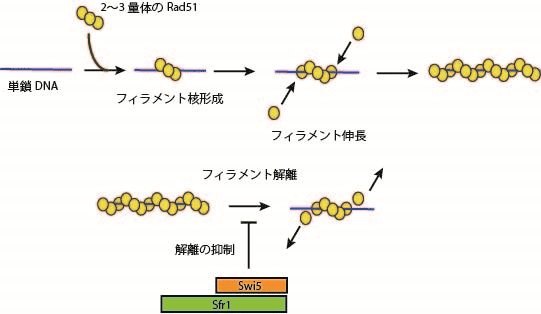
図1. Rad51-単鎖DNAフィラメント形成モデルとSwi5-Sfr1タンパク質による安定化
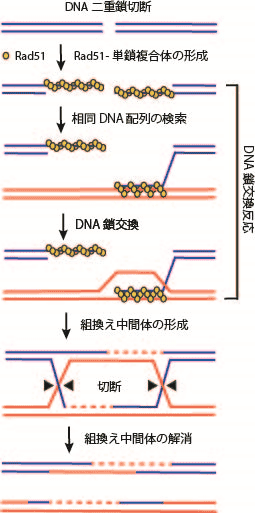
図2. 相同組換えによるDNA二重鎖切断の修復モデル
相同組換えは、減数分裂時に父方由来遺伝子と母方由来遺伝子をシャッフルして遺伝的多様性を創出する。一方で、DNAの損傷(特に、DNAの2本鎖が両方とも切断されるDNA二重鎖切断)の修復にも大きな役割を果たす。
DNAは紫外線や放射線など外的要因や、DNA複製の阻害や代謝で発生した活性酸素などの内的要因によって、日々常に損傷を受けており、相同組換えによるDNA修復が正常に働かない場合、ガンなどの重篤な疾患や不妊の原因になることが知られている。
相同組換えの中心的な反応は、DNA鎖交換反応であり、Rad51タンパク質によって触媒される。この反応は、Rad51が1本鎖DNAと結合しフィラメント状の複合体を形成し、複合体が二重鎖DNAを捕捉して似た配列を検索する。そして、似た配列を見つけるとDNA鎖を交換し、組換えを進行させる。DNA鎖交換反応において、Rad51と1本鎖DNAとの複合体形成は反応開始のスイッチとなる重要な過程で、様々な因子によって形成や解離が制御されている(図2)。
今回の研究では、Swi5-Sfr1によるRad51の1本鎖DNAへの結合・解離の様子を一分子かつリアルタイムで観察することで、DNA鎖交換反応開始に必須な複合体形成の分子機構の解明に成功した。
Rad51―単鎖DNAのフィラメント形成を制御する因子はSwi5-Sfr1タンパク質の他にも様々なものが存在することが知られている。有名なものでは、家族性乳ガンの原因遺伝子であるBRCA1、BRCA2などがある。これら因子がどのようにRad51―単鎖DNAフィラメントを制御するのか、より具体的な議論が可能となるばかりでなく、実際にこれらタンパク質を用いて解析ができれば、このガン抑制因子が、どのようにDNA鎖交換反応開始のスイッチとして働くのかという医学的に重要な分子機構に迫っていくことができる。
用語説明
[用語1] DNA―Rad51タンパク質 : 分子量約38,000のRad51タンパク質が数10~数100のヌクレオチドからなる1本鎖DNAに連なってらせん状に結合して、フィラメント状の高次DNA―タンパク質複合体を形成する。この複合体は中のDNA配列と相同な二重鎖DNAを検索し、相同な二重鎖DNAが見つかると鎖を交換する反応がおこる。
[用語2] リコンビナーゼ : 相同組換え(homologous recombination)において中心的な反応はRad51などのDNA鎖交換反応である。そのため、DNA鎖交換反応を促進するタンパク質がリコンビナーゼ(recombinase)と呼ばれるようになった。相同組換え以外のDNA組換え現象(例えば、部位特異的組換え)も知られており、その場合は、Rad51とは質的に全く異なる反応を触媒するリコンビナーゼが働く。そもそも、部位特異的組換えに働く酵素が最初に“リコンビナーゼ”と命名され、Rad51などはDNA鎖交換タンパク質と呼ばれていたが、21世紀以降、Rad51などもリコンビナーゼと呼ばれるようになった。
論文情報
掲載誌 : |
Proceedings of the National Academy of Sciences of the USA |
論文タイトル : |
Swi5–Sfr1 stimulates Rad51 recombinase filament assembly by modulating Rad51 dissociation |
著者 : |
Chih-Hao Lu, Hsin-Yi Yeh, Guan-Chin Su, Kentaro Ito, Yumiko Kurokawa, Hiroshi Iwasaki, Peter Chi, and Hung-Wen Li |
DOI : |
お問い合わせ先
東京工業大学 科学技術創成研究院 細胞制御工学研究センター 教授
岩﨑 博史(いわさき ひろし)
E-mail : hiwasaki@bio.titech.ac.jp
Tel : 03-5734-2588 / Fax : 03-5734-3781
取材申し込み先
東京工業大学 広報・社会連携本部 広報・地域連携部門
E-mail : media@jim.titech.ac.jp
Tel : 03-5734-2975 / Fax : 03-5734-3661
