東工大ニュース
東工大ニュース
![]()
公開日:2022.05.10
東京工業大学生命理工学院生命理工学系の西原秀典助教は、山梨大学の瀬川高弘講師、国立科学博物館の甲能直樹グループ長、東京農業大学の米澤隆弘准教授、国立遺伝学研究所、山形大学、国立歴史民俗博物館などとの研究グループにて、日本列島に生息していたオオカミの化石を用いてゲノムDNAの解析と放射性炭素による年代測定に成功しました。その結果、従来のニホンオオカミの起源に関する定説を覆し、更新世[用語1]の日本列島にはこれまで知られていない古い系統の大型オオカミが生息していたこと、またニホンオオカミの祖先は、更新世の古い系統のオオカミと最終氷期の後期に日本列島に入ってきた新しい系統の交雑により成立したことを初めて明らかにしました。本研究成果は日本時間の5月10日に米科学雑誌Current Biology(カレント・バイオロジー)電子版に掲載されました。
かつて日本には、極めて小型の日本固有亜種ニホンオオカミ(Canis lupus hodophilax)が生息していました。ニホンオオカミは古くは約9000年前の遺骸が見つかっており、本州・四国・九州に広く分布した肉食哺乳類でしたが、20世紀初頭に絶滅したことが知られています。一方、2万年前より以前の更新世の日本には世界最大級の巨大なオオカミが生息していたことが化石記録から知られていますが、このオオカミの系統は一切不明でした。そのため、更新世の巨大なオオカミと小型のニホンオオカミとの進化的な関連性については長年論争となっていました。そこでニホンオオカミの遺伝的起源を明らかにするために、3万5000年前の巨大な更新世オオカミの化石と5000年前のニホンオオカミの遺骸から古代DNA解析[用語2]を行いました。その結果、ミトコンドリアDNA[用語3]解析から更新世オオカミはニホンオオカミとは全く異なり古くに分岐した系統であることが明らかとなりました。また分岐年代推定[用語4]により、巨大な更新世オオカミは5万7000年前~3万5000年前の間に大陸から日本列島へ渡り、その後、3万7000年前~1万4000年前の間にニホンオオカミの祖先に繋がる系統が渡来したことが示されました。さらに核ゲノムDNA解析から、5000年前のニホンオオカミは巨大な更新世オオカミの系統と後から日本列島に入ってきた新しい系統が交雑して成立したことが明らかになりました(図1)。
本研究から、ニホンオオカミが従来考えられていたよりもはるかに複雑な進化史を持っていたこと、またその成立には日本列島という特殊な地理的環境が大きく寄与したことが初めて明らかになりました。
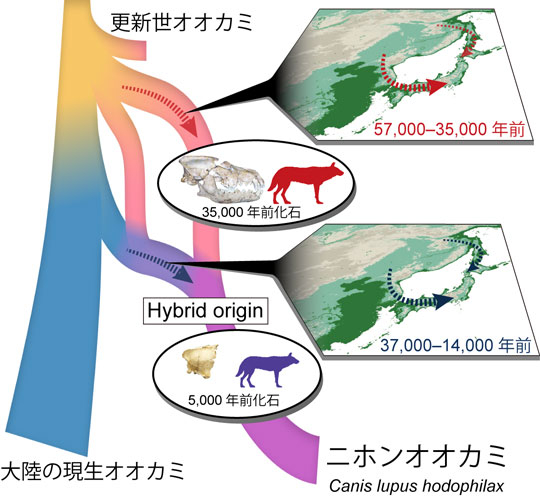
図1. 研究成果の概要図
ニホンオオカミ(Canis lupus hodophilax)は、日本列島にのみ生息していたハイイロオオカミ(Canis lupus)の絶滅した亜種です。ニホンオオカミは、オオカミ信仰としても知られるように古来より日本人に畏敬の念をもたれてきた動物でしたが、1905年に確認されたのを最後に絶滅しました。1800年代前半、ドイツの医師で博物学者のシーボルトはニホンオオカミの剥製標本をオランダの動物学者テミンクに送りました。テミンクが1842年に出版したFauna Japonica(日本動物誌:初の日本の動物学に関する一連の刊行物)にニホンオオカミを記載したことにより、日本固有であるニホンオオカミの存在が広く世界に知られました。
ニホンオオカミは、平均頭骨長が約196 mm、歯(下顎第一大臼歯, m1)の長さが約24 mmと、現存ならびに絶滅した世界中のハイイロオオカミの中でも極めて小柄な亜種です。これは島嶼化[用語5]の結果であると考えられていますが、このニホンオオカミの進化史はほとんど分かっておらず大きな未解決問題の1つです。ミトコンドリアDNAの研究から、小柄なニホンオオカミは現存する世界のハイイロオオカミの中で最も早く分岐した系統の1つであることが示唆されています。また、ニホンオオカミの最古の遺骸は9000年前まで遡ることができるため、それ以前にニホンオオカミの祖先がユーラシア大陸から日本列島に移動してきたと考えられています。なお、北海道ではかつてハイイロオオカミの亜種であるエゾオオカミ(Canis lupus hattai)が生息していましたが、形態学的ならびにDNAの分析からニホンオオカミとは別の系統であることが既に知られています。
一方で、2万年より前には世界最大級のオオカミが日本列島に生息していたことが分かっています(図2)。この本州の更新世オオカミの歯(下顎第一大臼歯, m1)は、最大で34.5 mmにも達します。現生のハイイロオオカミのm1が24.0~33.5 mm、大陸の更新世のオオカミ化石でも28.1~33.4 mmの範囲であることから、日本列島の更新世オオカミは地史的な記録における全てのハイイロオオカミの中で最大級であったことが分かっています。
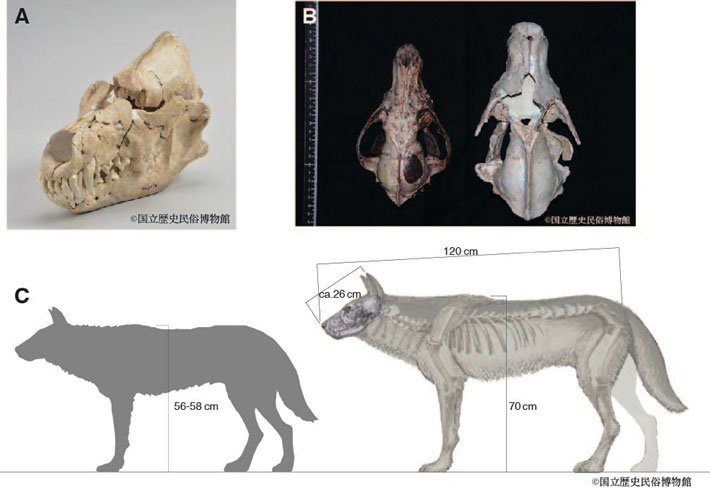
図2. 本研究で分析した本州に生息していた更新世オオカミとニホンオオカミ
(A)今回分析した栃木県産の更新世オオカミの頭骨
(B)神奈川県で捕獲された標準的なニホンオオカミの標本(Canis lupus hodophilax)(左)と更新世オオカミ(右)との頭骨サイズの比較
(C)ニホンオオカミ(左)と更新世オオカミ(右)の体格の差異
この世界最大級の更新世オオカミの系統的位置づけや、小柄なニホンオオカミとの関係は一切不明であり、長年の謎とされてきました。これまで主に次の2つの仮説(1)巨大な更新世オオカミはニホンオオカミの直接の祖先であり、この更新世オオカミが島嶼適応によって小型化を遂げてニホンオオカミとなった。(2)巨大な更新世オオカミとニホンオオカミは別種である。が提唱されていました。しかしながら、化石の形態のみでは種内の進化史の実態に迫るのは困難であり、長年論争が続いていました。したがって、ニホンオオカミの起源と進化史を理解するためには、巨大な更新世オオカミのDNA情報に基づいた解析が必要でした。
過去の動物や古人骨からDNAを分析する古代DNA研究は、過去の動物や人類の系統や進化を知る上でとても有力な方法です。しかし、日本は高温多湿かつ酸性土壌が多く、化石に残存しているDNAの保存状態が極めて悪い環境にあります。そのため技術的な難しさから、更新世オオカミの古代DNA解析は全く行われていませんでした。本研究では、最先端の古代DNA解析技術を用いて、3万5000年前の巨大な更新世オオカミおよび5000年前の縄文時代のニホンオオカミ標本から古代 DNA解析を行いました。これは日本国内最古の試料からの古代DNA研究になります。
栃木県佐野市から産出したオオカミ化石標本の内部から試料を採取し、放射性炭素年代測定、炭素・窒素の安定同位体比の測定、ならびにDNA抽出と次世代シーケンサー[用語6]による塩基配列の解読を行いました。放射性炭素年代測定からは化石の年代が3万5000年前(後期更新世)と5000年前(完新世)であることが明らかになりました。また安定同位体比の分析から、3万5000年前の更新世オオカミは5000年前のニホンオオカミと比較して、炭素と窒素の安定同位体比が高い値を示したことから、巨大な更新世オオカミはニホンオオカミよりも強い肉食性で大型草食哺乳類を捕食していたことが推察されました。
また3万5000年前の大型の更新世オオカミから得られたミトコンドリアDNA情報について系統解析[用語7]を行いました。その結果、これまで本州・四国・九州には絶滅したニホンオオカミの1亜種しか生息していなかったと認識されていましたが、この更新世オオカミは既知のニホンオオカミとは全く異なる系統であり、現生のオオカミ系統よりも古くに分岐した系統であることが示されました(図3)。絶滅した古い系統である更新世オオカミは、これまでシベリアやアラスカなどの亜寒帯などからしか報告がありませんでしたが、その更新世オオカミ系統の一部が日本列島にまで到達していたことが初めて明らかになりました。
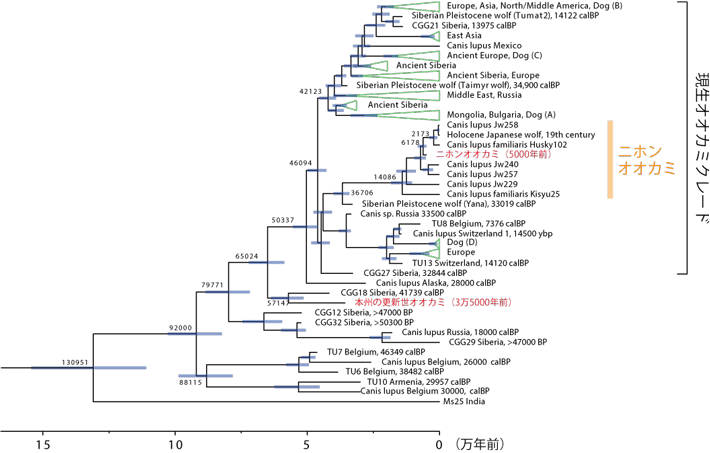
図3. ミトコンドリアDNA解析に基づくハイイロオオカミの系統関係と分岐年代推定
系統樹上の数値は推定分岐年代の中央値。本研究で用いた2個体を赤字で示す。5000年前のニホンオオカミは既知のニホンオオカミグループに含まれ、3万5000年前の更新世オオカミは現生オオカミのグループよりも古くに分岐したことが示された。
さらにより詳細なオオカミ集団の歴史を調べるために、5000年前のニホンオオカミと3万5000年前の更新世オオカミの核ゲノム配列を現生のオオカミや絶滅した更新世の大陸のオオカミとともに解析しました。その結果、ニホンオオカミは、3万5000年前の日本の更新世オオカミと大陸のオオカミの双方から遺伝的な寄与を受けて成立したという強い証拠が見つかりました(図4)。ミトコンドリアDNAではニホンオオカミが大陸の現生オオカミの系統に含まれることが示されましたが(図3)、これは現生オオカミの祖先に近い系統からの遺伝子の流入を反映した結果と考えられます。この結果から、ニホンオオカミが複数の系統の交雑によって成立したことが明らかになりました。
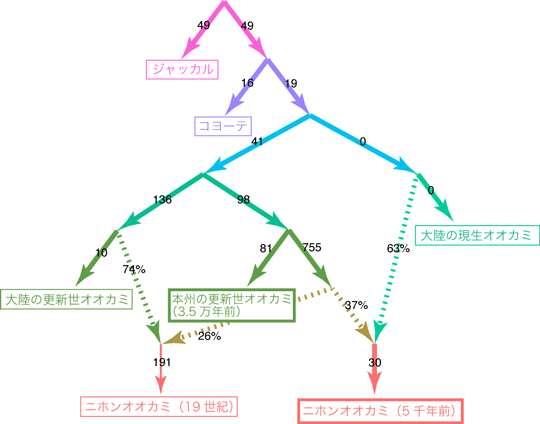
図4. 核ゲノムデータに基づく更新世オオカミ集団と大陸の現生オオカミ集団の関係と交雑の歴史
3万5000年前の日本の更新世オオカミの集団と大陸のオオカミに由来する集団の交雑により、ニホンオオカミが成立した。
今回の結果から考えられるニホンオオカミの進化的起源は次のようになります(図5)。まず大陸に生息していた更新世オオカミ系統の1つが5万7000年前~3万5000年前の間に日本列島へ渡り、ユーラシア大陸とは異なる孤立した系統を形成していました。日本列島で発見されている巨大な更新世オオカミの化石がこれにあたります。その後3万7000年前~1万4000年前の間に、現生オオカミの祖先やシベリアの一部の更新世オオカミと遺伝的交流を持つオオカミが日本列島へ渡りました。日本列島では少なくともこの2つの系統が交雑することでニホンオオカミが成立し、その後大陸と隔離された地理的環境下で繁栄したと考えられます。このようにニホンオオカミは複雑な遺伝的起源によって極めてユニークなゲノム情報を持つに至ったことが明らかになりました。
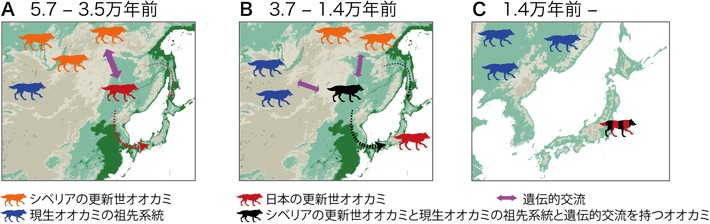
図5. 本研究結果から推定されたニホンオオカミの起源
(A)日本の更新世オオカミは5万7000年前~3万5000年前の間に日本列島に移動した。
(B)シベリアの更新世オオカミと現生オオカミの祖先系統と遺伝的交流を持つオオカミが、約3万7000年前~1万4000年前の間に日本列島に移動した。
(C)交雑の結果、複数のオオカミ系統のゲノムを持つニホンオオカミが成立した。最終氷期以降、日本列島のオオカミは大陸のオオカミから地理的に完全に隔離され、本州・四国・九州で生存していたが、1900年代初頭に絶滅した。 色の異なるオオカミは異なる集団を示す。破線の矢印は、北または南からの移動経路の可能性を示している。表示されている日本地図は、最終氷期最盛期の海や海岸線の地形を反映している。地図の緑の部分は大陸氷塊の蓄積により海面が120 m低下した場合の陸域を示している。
巨大な更新世オオカミがニホンオオカミの直接の祖先であるか否かについては、長年の論争がありました。本研究から判明した更新世オオカミとニホンオオカミの関係性は、これまでのどちらの仮説とも異なるものであり、ニホンオオカミの進化史が極めて特殊であることを示しました。しかしニホンオオカミがなぜ小型でありそれにはどのような遺伝的要因があったのかについてはまだ課題が残されています。これを解明するにはさらに多くのニホンオオカミ、特に更新世や縄文時代など古い年代のオオカミ化石の古代DNA解析を進めることが重要になります。
日本列島は基本的にユーラシア大陸から隔離された島弧でありながら、氷期の海水準低下期に断続的に陸続きになるという特殊な地理的環境です。こうした環境であったからこそ、今回の成果は大陸集団では検出できない時間軸を伴った生物地理学的な挙動まで解明できたと言えます。後期更新世以前(1万2000年以前)の日本列島にはヒグマ、バイソン、オオツノジカ、ヘラジカ、トラ、ナウマンゾウといった様々な大型哺乳類が生息していました。それらは大陸から断続的に渡来した結果、「重層的」に集団の置き換わりが起こった歴史を持つ可能性があり、今後の古代DNA研究の最も重要なテーマの1つになると考えられます。実際、更新世の日本列島に生息していたヒグマもユーラシア大陸から日本列島への複数回の移動の歴史があったことが最近明らかになっています。しかも本研究で示されたように、古い系統が新しい系統に完全に置き換わるのではなく交雑することで新しい集団が形成されていくという過程は、ニホンオオカミのみならず普遍的に起こってきた可能性があり、今後日本列島における哺乳類相の進化史の解明が進むと期待されます。また本研究で使用した微量DNA分析技術は様々な分野に応用可能であり、今後は様々な年代の古い試料や損傷した試料からのDNA分析に応用したいと考えています。
研究体制と支援
本研究は、山梨大学の瀬川高弘講師、国立科学博物館の甲能直樹グループ長、東京農業大学の米澤隆弘准教授、東京工業大学の西原秀典助教、国立遺伝学研究所の森宙史准教授、国立極地研究所の秋好歩美学術支援技術専門員、東海大学の呉佳斎研究員、国立科学博物館の甲能純子協力研究員、学習院女子大学の工藤雄一郎准教授、山形大学の門叶冬樹教授、国立歴史民俗博物館の坂本稔教授からなる研究チームによって行われました。
本研究は、日本学術振興会科研費(課題番号: 20K20942)、国立科学博物館の化学層序プロジェクトと国立歴史民俗博物館の直良信夫コレクション共同研究助成の支援を受けました。
用語説明
[用語1] 更新世 : 地質時代の区分の1つで、約258万年前~約1万2千年前までの期間。更新世は細かい時代に分けられ、その最後の時代である後期更新世が約13万年前~1万2千年前。
[用語2] 古代DNA解析 : 化石など古い時代の生物に由来する標本に残された微量のDNA配列を解析する手法。近年の古代DNA解析技術の発展により様々な絶滅生物や古人類の進化系統解析に用いられるようになってきた。
[用語3] ミトコンドリアDNA : 細胞内小器官のミトコンドリアが独自に持つDNAであり、ミトコンドリアDNAは母親から子へ母系遺伝する。塩基置換率が高いことから、系統解析および分岐年代推定によく用いられる。
[用語4] 分岐年代推定 : 生物間のDNA配列あるいはアミノ酸配列の違いを比較して、系統樹上の分岐した年代を推定する方法。
[用語5] 島嶼化 : 大陸から隔離された島嶼部で見られる進化の傾向に関する説。島嶼部特有の生態系や資源に応じて動物の体サイズが巨大化もしくは矮小化すると考えられている。
[用語6] 次世代シーケンサー : 数億に及ぶ DNA断片配列を高速に決定できる装置。この機器を用いることにより古い化石標本に残された微量DNAの配列を決定することが可能となる。
[用語7] 系統解析 : 塩基配列やアミノ酸配列の違いを統計的に比較解析することで、生物種間あるいは個体間の類縁関係を推定する解析のこと。
論文情報
掲載誌 : |
Current Biology |
論文タイトル : |
Paleogenomics reveals independent and hybrid origins of two morphologically distinct wolf lineages endemic to Japan. (古代DNA解析が明らかにしたニホンオオカミの起源の解明) |
著者 : |
Takahiro Segawa, Takahiro Yonezawa, Hiroshi Mori, Ayako Kohno, Yuichiro Kudo, Ayumi Akiyoshi, Jiaqi Wu, Fuyuki Tokanai, Minoru Sakamoto, Naoki Kohno, Hidenori Nishihara |
DOI : |
お問い合わせ先
山梨大学 総合分析実験センター
瀬川高弘
E-mail : tsegawa@yamanashi.ac.jp
Tel : 055-273-9439
国立科学博物館 地学研究部
甲能直樹
E-mail : kohno@kahaku.go.jp
Tel : 029-853-8196
東京農業大学 農学部
米澤隆弘
E-mail : ty205685@nodai.ac.jp
Tel : 0462-70-6572
東京工業大学 生命理工学院
西原秀典
E-mail : hnishiha@bio.titech.ac.jp
取材申し込み先
山梨大学 企画部広報企画課広報企画グループ
E-mail : koho@yamanashi.ac.jp
Tel : 055-220-8006 / Fax : 055-220-8799
国立科学博物館 広報・運営戦略課広報戦略担当
E-mail : t-shuzai@kahaku.go.jp
Tel : 029-853-8984 / Fax : 029-853-8998
東京農業大学 経営企画部
E-mail : koho@nodai.ac.jp
Tel : 03-5477-2300 / Fax : 03-5477-2707
東京工業大学 総務部 広報課
E-mail : media@jim.titech.ac.jp
Tel : 03-5734-2975 / Fax : 03-5734-3661
国立遺伝学研究所 リサーチ・アドミニストレーター室広報チーム
E-mail : prkoho@nig.ac.jp
Tel : 055-981-5873 / Fax : 055-981-9418
山形大学 総務部総務課秘書広報室
E-mail : yu-koho@jm.kj.yamagata-u.ac.jp
Tel : 023-628-4008 / Fax : 023-628-4013
資料に関する問い合わせ
国立歴史民俗博物館 広報サービス室広報・普及係
E-mail : koho@ml.rekihaku.ac.jp
Tel : 043-486-6488(直通) / Fax : 043-486-4482