東工大ニュース
東工大ニュース
![]()
公開日:2019.08.08
東京工業大学バイオ研究基盤支援総合センターの廣田順二准教授と榎本孝幸研究員(研究当時)らは、生命理工学院の梶谷嶺助教、伊藤武彦教授、米国モネル化学感覚研究所の松本一朗博士らと共同で、鼻の中でタイプの異なる2種類の匂いセンサー細胞(嗅神経細胞)が作り出されるメカニズムを世界で初めて明らかにした。
様々な匂い物質を感知する嗅覚受容体はゲノム上で最大の遺伝子ファミリー[用語1]を形成している。嗅覚受容体ファミリーは、魚類から哺乳類に至る脊椎動物に共通して存在するクラスI(水棲型)と陸棲動物に特異的なクラスII(陸棲型)の2種類に大きく分類される。つまり鼻の中では発現する受容体のタイプによって異なる2種類の嗅神経細胞が作り出されるが、そのメカニズムは長年未解明のままだった。
今回研究グループは、2種類の嗅神経細胞[用語2]を作り分けるのに必要な転写因子[用語3]を発見し、その生体内での機能を明らかにして、2タイプの嗅神経細胞ができる仕組みを解明した。さらに、この転写因子の発現を人為的に操作するだけで、鼻の中が主に水棲型の嗅神経細胞もしくは陸棲型の嗅神経細胞からなるマウスの作出に成功した。興味深いことに、水棲型と陸棲型のバランスが崩れたマウスは、例えば、食べ物の腐った臭いに極端に敏感になる一方、天敵の臭いをあまり嫌がらなくなるなど、感じとる「匂いの世界」が大きく変化することがわかった。
研究成果はSpringer Nature(シュプリンガー・ネイチャー)社の科学誌『Communications Biology』(オンライン版)に8月7日に公開された。
嗅覚は環境中の匂い物質を感知し、食物探索・天敵からの回避・生殖など個体の生存や種の保存に必要な行動を誘導する重要な感覚である。多種多様な匂い物質を受容する嗅覚受容体はゲノム上最大の遺伝子ファミリーを形成し、その数はマウスで約1,100個にも及び、実にマウスの全遺伝子の約5%を占める(ヒトでは約400個、ヒト全遺伝子の約2%)。
この巨大な嗅覚受容体ファミリーは、大きくクラスI型とクラスII型の2種類に分類される。クラスIは「魚類から哺乳類に共通したタイプ(水棲型)」で水に溶けやすい親水性の匂い分子を、クラスIIは「陸棲動物特異的なタイプ(陸棲型)」でより水に溶けにくい疎水性の高い匂い分子を受容すると考えられている。
両者は神経解剖学的にも明確に区別される。クラスI嗅覚受容体を発現する嗅神経細胞(クラスI嗅神経細胞)は嗅上皮[用語4]の背側に限局して存在し、嗅球(脳の一部)の背側領域に神経接続する。一方、クラスII嗅神経細胞は嗅上皮全体に分布するが、嗅球のクラスIより腹側の領域に接続する。
嗅神経細胞が作り出される過程では、細胞はまずクラスIかクラスIIかのいずれかの運命を選択し、その運命決定に従い、対応するクラスの嗅覚受容体レパートリーから、1種類の受容体のみを選択的に発現する。こうして嗅神経細胞は発現する嗅覚受容体が異なる多様な個性を有する細胞集団として産生される。
この多様性こそが複雑な匂い情報の感知・識別を可能にする分子基盤を形作っている。リンダ・バック(Linda・Buck)とリチャード・アクセル(Richard・Axel)の嗅覚受容体の発見(2004年ノーベル生理学医学賞)以降、嗅神経細胞における「1細胞1受容体」の発現機構の解明が大きく進展してきた一方で、その前段階である「嗅覚受容体のクラス選択」は嗅覚機能の基盤をなす重要なプロセスであるにもかかわらず、その分子メカニズムは長年未解明のままだった。
細胞の運命決定は転写因子と呼ばれるタンパク質によって制御されることが知られている。今回、研究グループは「嗅覚受容体のクラス選択は、クラス特異的に発現する転写因子によって制御される」との仮説のもと、トランスクリプトーム解析[用語5]を通じてクラスII嗅神経細胞のみに発現する転写因子Bcl11bを同定した。
この転写因子Bcl11bの機能を明らかにするために、マウスにおいてBcl11b遺伝子を欠損させたところ、変異マウスの鼻の中ではクラスI嗅神経細胞の数が大幅に増加し、一方クラスII嗅神経細胞の数が激減していることを見出した(図1)。逆にすべての嗅神経細胞にBcl11bを強制的に発現させたところ、今度はクラスI嗅神経細胞がほぼ消失してしまった。
つまり嗅神経細胞は、Bcl11b非存在下ではクラスIの運命を選択し、Bcl11b存在下ではクラスIの運命選択は抑制され、クラスIIの運命選択が可能になることが示された。この一連の実験により、嗅覚受容体のクラス選択を制御する転写因子がBcl11bであることを世界で初めて明らかにした。
では、Bcl11bがクラスIの運命選択(クラスI嗅覚受容体の発現)をどのように抑制しているのか。今回の研究は、その分子メカニズムも解明した。ゲノム上に長大な単一の遺伝子クラスター[用語6]を形成するクラスI嗅覚受容体遺伝子の発現は、J-エレメント[用語7]と名付けられた超長距離作用性のエンハンサー[用語8]によって制御されることが同研究グループによって明らかにされている(Iwata et al., Nature Communications 2017)。今回の研究のBcl11bとJ-エレメントの機能相関を明らかにする実験から、Bcl11bがJ-エレメントのエンハンサー活性を負に制御することで、クラスI嗅覚受容体の全体の遺伝子発現を抑制していることが明らかになった。
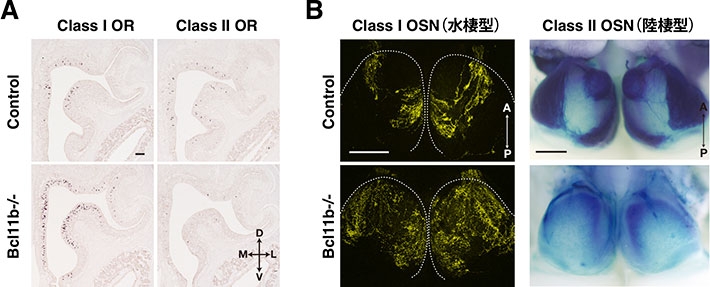
図1. 転写因子Bcl11bによる嗅覚受容体のクラス選択の制御
(A)転写因子Bcl11bを欠損したマウス(Bcl11b-/-)の嗅上皮では、嗅神経細胞の多くがClass I嗅覚受容体(Class I OR)を発現し、Class II嗅覚受容体(Class II OR)の発現は激減する。
(B)嗅神経細胞の脳(嗅球)への投射領域はBcl11bの欠損によって大きく変化する。コントロールでは、Class I嗅神経細胞は嗅球の最も背側の特定の領域(黄色の蛍光)に投射し、Class II嗅神経細胞はそれを避けた部分(青色の染色)に投射する。Bcl11b欠損によって水棲型の鼻になったマウス(Bcl11b-/-)では、嗅球のほとんどがClass I嗅神経細胞(Class I OSN)で支配され、Class II嗅神経細胞(Class II OSN)の軸索投射はほとんど認められなくなる。
研究グループは嗅覚受容体のクラス選択の分子メカニズムを明らかにしたことで、人為的に転写因子Bcl11bの発現を制御することで嗅神経細胞の運命をコントロールできることを示した。その結果、主にクラスI嗅神経細胞からなる水棲型の鼻をもったマウス(図1)と、主にクラスII嗅神経細胞からなる陸棲型の鼻をもったマウスを作り出すに成功した。
これら遺伝子改変マウスが感じる「匂いの世界」がどのように変わるのかを明らかにするために、2種類の匂い物質を用いて、マウスの匂いに対する行動を解析した。用いた匂い物質は、クラスI嗅覚受容体が感知するとされる腐敗臭の原因物質2-メチル酢酸、もう1つはクラスII嗅覚受容体が感知すると考えられている天敵臭TMT(トリメチルチアゾリン)である。
野生型マウスは両方の臭いを嫌がる行動を示すが、興味深いことに、水棲型の鼻をもったマウスは腐敗臭を極端に嫌がる一方で、天敵臭をあまり嫌がらなくなった(図2)。逆に陸棲型の鼻のマウスは、腐敗臭をあまり嫌がらなくなり、天敵臭は野生型と同様に嫌がることがわかった。つまり鼻の中の異なる2つのタイプの嗅神経細胞のバランスが崩れることで、マウスが感じる匂いの世界は大きく変わることがわかった。

図2. Bcl11bの機能欠損変異と機能獲得変異によってマウスが感じる匂いの世界は大きく変化
通常、野生型のマウスの鼻はClass I(水棲型)が15%、Class II(陸棲型)が85%を占める。Bcl11b機能欠損変異(Bcl11b cKO)マウスではほぼClass I嗅神経細胞が占める水棲型の鼻に、一方、Bcl11b機能獲得変異(Bcl11b Overexpression)マウスの鼻からはClass I嗅神経細胞がほぼ消失し、陸棲型(Class II)の鼻となる。水棲型の鼻をもったマウスは、腐敗臭(2MBA)を極度に嫌がり、壁をよじ登って逃げようとする(点線)一方で、天敵臭(TMT)はあまり嫌がらなくなる。陸棲型の鼻をもつマウスは腐敗臭をあまり嫌がらなくなってしまう。
比較的変化の少ない水中の環境と比べて、陸上の環境は多種多様で変化に富んでいる。水棲から陸棲への進化の過程で動物を取り巻く環境は大きく変化し、その結果クラスII(陸棲型)嗅覚受容体が陸棲動物特異的にその数を爆発的に増やし、分子進化したと考えられている。すなわち嗅覚の陸棲適応である。
今回解明したBcl11bによる嗅覚受容体のクラス選択の分子メカニズムは、嗅覚の陸棲適応時に動物が獲得した可能性が考えられる。この仮説を検証するために、単一の個体で水棲から陸棲へと生活環境を変える両生類のカエルに着目した。これまでの研究から、水中で生活するオタマジャクシの鼻にはクラスI(水棲型)嗅覚受容体のみが発現している。成体のカエルの鼻では、空中にでる部分(air nose)にクラスII(陸棲型)嗅覚受容体が、水に浸かっている部分(water nose)にクラスI(水棲型)嗅覚受容体が発現する。
転写因子Bcl11bのカエルでの発現パターンを解析したところ、水中で生活するオタマジャクシの嗅神経細胞ではBcl11bは発現しておらず、変態期になって初めて将来air noseとなる部分でBcl11bが発現し始め、成体となったカエルではクラスII嗅神経細胞が存在するair noseにのみにBcl11bが発現していることがわかった(図3)。これは、Bcl11bが嗅覚の陸棲適応に深く関わっていることを示唆する結果であり、今回の研究成果の生物学的、進化学的重要性を示すものとなっている。
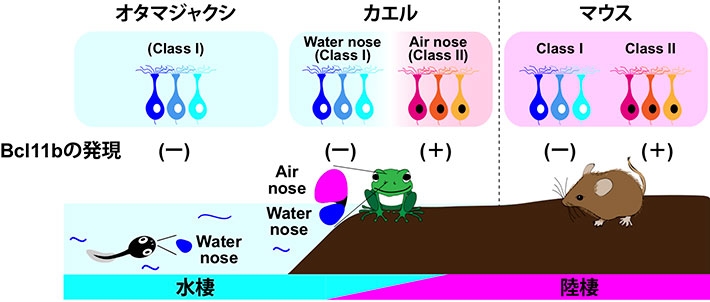
図3. Bcl11bによる嗅覚受容体クラス選択と嗅覚の陸棲適応モデル
転写因子Bcl11bはClass II嗅神経細胞特異的に発現する。本研究によってBcl11bは水棲型のClass I嗅覚受容体の発現を抑制することで、陸棲型のClass II嗅覚受容体の発現を可能にすることが明らかになった。Class II嗅覚受容体は陸棲動物特異的な受容体であることから、この分子メカニズムは嗅覚の陸棲適応を解く鍵になると考えられる。実際、オタマジャクシ(水棲)からカエル(両棲)への生活環境の変化に伴い、Bcl11bは空中にでる鼻においてClass II嗅覚受容体と共発現しており、嗅覚の陸棲適応との相関が認められた。
研究グループは、嗅覚受容体の発見以来、四半世紀以上未解明であった嗅覚受容体のクラス選択の分子メカニズムを明らかにした。さらに末梢神経である嗅神経細胞の運命を変えるだけでマウスが感じる匂いの世界と嗅覚行動は大きく変化することを示した。しかし、嗅覚受容体のクラス選択、つまり末梢から中枢への感覚入力の変化が、脳内の神経回路形成や情報処理に及ぼす影響はブラックボックスのままである。
今後、Bcl11bによる嗅覚受容体のクラス選択の遺伝学的操作を通じ、これらを明らかにすることで、嗅覚受容体クラス選択と動物の嗅覚行動を繋ぐ神経基盤が解明できるものと期待される。
論文情報
掲載誌 : |
Communications Biology |
論文タイトル : |
Bcl11b controls odorant receptor class choice in mice |
著者 : |
Takayuki Enomoto, Hidefumi Nishida, Tetsuo Iwata, Akito Fujita, Kanako Nakayama, Takahiro Kashiwagi, Yasue Hatanaka, Hiro Kondo, Rei Kajitani, Takehiko Itoh, Makoto Ohmoto, Ichiro Matsumoto, and Junji Hirota* |
DOI : |
用語説明
[用語1] 遺伝子ファミリー : 進化上同一の祖先遺伝子に由来すると考えられる、配列や機能が類似した遺伝子群。嗅覚受容体遺伝子ファミリーは、7回膜貫通型Gタンパク質共役型受容体という特徴を共有する巨大ファミリーである。
[用語2] 嗅神経細胞 : 鼻腔内で匂いを感知する器官(嗅上皮)に存在し、匂い物質の感知に特化した神経細胞。一つの嗅神経細胞は膨大な嗅覚受容体ファミリーから一種類のみを選択的に発現する。
[用語3] 転写因子 : ゲノムDNA上の特定の塩基配列に結合するタンパク質の総称。転写因子は、DNAが有する遺伝情報の読み出しを促進、または逆に抑制し、遺伝子発現を調節する。
[用語4] 嗅上皮 : 鼻腔の上部にある嗅覚器官。匂いを感知する嗅神経細胞があり、粘膜に覆われている。
[用語5] トランスクリプトーム解析 : 特定の細胞や組織で発現している遺伝子を網羅的に解析すること。数千から数万の遺伝子を一度にプロファイリングして、細胞機能の全体像を把握するための解析。
[用語6] 遺伝子クラスター : 同様の機能を有する多数の遺伝子が、染色体上の同じ位置に直列して位置している状態(遺伝子集団)。クラスI嗅覚受容体遺伝子クラスターは、129個の遺伝子が約300万塩基対の範囲内に密集して存在する、極めて巨大な遺伝子クラスターの1つ。
[用語7] J-エレメント : クラスI嗅覚受容体遺伝子の発現を制御する転写調節領域(エンハンサー)。J-エレメントはクラスI嗅覚受容体クラスター全体を制御し、制御する遺伝子の数とゲノム上の距離において、これまでに類をみない規模で遺伝子発現を調節する。
[用語8] エンハンサー : ゲノム上には、タンパク質をコードする遺伝子領域のほかにも様々な機能を持つ配列が存在する。その一つである「エンハンサー」は、遺伝子発現を促進する配列として、遺伝子が発現するタイミングや組織を規定する。
本研究は文部科学省科研費「新学術領域研究」(JP21200010、18H04610、19H05256)、日本学術振興会科研費「基盤研究C」(JP20570208、JP16K07366)、「基盤研究B」(JP19H03264)、「若手研究B」(JP25840085、JP16K18361、17K14932)、千里ライフサイエンス財団、稲森財団、住友財団、倉田記念日立科学技術財団の支援を受けて行われた。
お問い合わせ先
東京工業大学 バイオ研究基盤支援総合センター
准教授 廣田順二
E-mail : jhirota@bio.titech.ac.jp
Tel : 045-924-5830 / Fax : 045-924-5832
取材申し込み先
東京工業大学 広報・社会連携本部 広報・地域連携部門
E-mail : media@jim.titech.ac.jp
Tel : 03-5734-2975 / Fax : 03-5734-3661